Способность фототрофных бактерий к фотосинтезу, как и у растений, определяется наличием магнийсодержащих порфириновых пигментов — хлорофиллов. Состав бактериальных хлорофиллов, называемых бактериохлорофиллами, отличается от хлорофиллов растений.
Свет в качестве первичного источника энергии могут использовать три главные группы бактерий - зеленые и пурпурные серные бактерии (Chlorobiaceae и Chromatiaceae) и. пурпурные несерные бактерии (Rhodospirillaceae). Как правило, фотосинтезирующие бактерии имеют клеточную стенку, сходную со стенкой других грамотрицательных бактерий, однако под этой стенкой у них расположена толстая клеточная мембрана, которая может многократно изгибаться и впячиваться, образуя ламеллярные мембранные структуры или везикулы внутри клетки. Эти структуры несут фотосинтетический аппарат, который значительно отличается от соответствующего аппарата хлоропластов растений.
В деталях биохимия фотосинтетических процессов у бактерий и растений также различна. Препараты фотосинтетических, мембран можно получить из разрушенных клеток в виде частиц или пузырьков, называемых хроматофорами, но, как правило, интактные клетки, за исключением клеток зеленых серных бактерий (Chlorobiaceae), не содержат таких дискретных структур. Зеленые серные бактерии (например, Chlorobium) отличаются от других: фотосинтезирующих бактерий тем, что лишены внутренних мембран; вместе с тем они обладают уплощенными сигарообразными структурами, называемыми хлоросомами, которые присоединены к плазматической мембране.
Хлоросомы служат функционально организованными светособирающими антеннами: для реакционных центров, которые локализованы в плазматической мембране. Перенос энергии в хлоросоме происходит с бактериохлорофилла с (1000-1500 молекул) через бактерио-хлорофилл а (возможно, 100 молекул в комплексе с белком) на бактерио-хлорофилл с реакционного центра.
Бактериальный фотосинтез во многом отличается от фотосинтеза у растений. Во-первых, бактерии - единственные из фотосинтезирующих организмов - не способны использовать в качестве конечного восстановителя воду. Они используют другие восстановители, которыми могут быть органические молекулы или неорганические соединения серы, и, следовательно, бактерии не выделяют кислород.
Во-вторых, фиксация и метаболизм углерода у бактерий происходят не в цикле Кальвина - Бенсона, а иным путем.
В-третьих, аппарат первичного улавливания света и переноса электронов у них совершенно отличен от наблюдающихся в растительных клетках; в частности, у них протекает только одна световая реакция, правда, она во многом сходна с реакцией в фотосистеме I растений.
Вместе с тем механизм улавливания света у бактерий очень сходен с соответствующим механизмом у растений, хотя фотосинтетические единицы у первых меньше. Так же как в хлоропластах, свет поглощается пигментами антенны, энергия возбуждения быстро передается на реакционный центр и используется в качестве движущей силы в транспорте электронов.
Главным фотоактивным пигментом является бактериохлорофилл (БХл), в большинстве случаев бактериохлорофилл а (10.15), а в некоторых случаях (например, у бактерии Rhodopseudomonas sphaeroides) - бактериохлорофилл b.
В тесной связи с реакционным центром находится главная светособирающая антенна, которая у Rhodospirillaceae поглощает при 875 нм. Этот комплекс включает два полипептида, а также бактериохлорофилл и каротиноиды в соотношении 1 : 1. Соотношение БХл: реакционные центры остается постоянным и составляет около 25: 1. У некоторых видов, например у Rhodospirillum rubrum, имеется только один светособирающий комплекс, тогда как у других организмов, например, у Rhodopseudotnonas spp., содержится и второй подобный комплекс, поглощающий при более коротких длинах волн (800 и 850 нм). Он включает два пептида, две формы бактериохлорофилла - (две молекулы БХл-850 и одну молекулу БХл-800) и одну молекулу каротиноида. Отношение числа этих комплексов к числу реакционных центров варьирует в зависимости от условий окружающей среды от 10 до 100 молекул БХл800-850 на один реакционный центр.
В первичной световой реакции энергия возбуждения переносится на Р-870 и один электрон передается от бактериохлорофилла "особой пары" на акцептор. Окисленный Р-870 в свою очередь получает электрон от молекулы донора. Химическая природа этого донора и первичного акцептора пока неизвестна.
В заключение следует отметить, что электронтранспортныс цепи у разных видов бактерий различаются в деталях. Наиболее подробно учёные их изучали у Rhodospirillum rubrutn, у нескольких видов Rhodopscudomonas (R. sphaeroides, R. capsulata, R. palustris) из Rhodospiriilaceae и у некоторых видов Chromatiutn и Chlorobium из классов серных бактерий. У всех этих бактерий в цепи переноса электронов участвует несколько цитохромов, тогда как плаетоцианин, по-видимому, не используется. Те функции, которые иластоцианин выполняет у растений, у бактерий может выполнять убихинон.
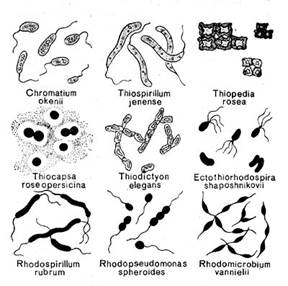
Фототрофные, или фотосинтезирующие, бактерии — типично водные микроорганизмы, распространенные в пресных и соленых водоемах. Особенно часто они встречаются в местах, где есть сероводород, как в мелководье, так и на значительной глубине. В почве фототрофных бактерий мало, но при затоплении ее водой они могут расти весьма интенсивно. Развитие фототрофных бактерий нередко легко обнаружить, не прибегая к постановке накопительных культур и микроскопическим исследованиям, так как многие из них способны образовывать ярко окрашенные пленки, а также обрастать подводные предметы. Такие макроскопические скопления наблюдаются в серных источниках, лиманах, бухтах, озерах и прудах. Иногда в результате массового развития фототрофных бактерий меняется даже цвет всей воды в водоеме или отдельные ее слои становятся окрашенными. Последнее явление довольно часто имеет место в некоторых озерах, содержащих в придонных слоях сероводород.
Поможем написать любую работу на аналогичную тему