Дыхание у микробов рассматривается как энергетический процесс или как совокупность различных химических реакций, окисления и расщепления веществ органического и неорганического происхождения. В результате этих химических реакций освобождается энергия, которая используется микробами для усвоения питательных веществ, синтеза бел кон их тела, движения, роста, размножения и других отправлений живого организма.
Примером высвобождения энергии может служить окисление глюкозы, которое можно выразить следующим у соединением:
С6 Н2О + 6 О 2 6 Н2О+ 6СО2 + 674 ккал.
Как видно из уравнений, в результате полного окисления одной молекулы глюкозы до конечных продуктов (воды и углекислого газа) выделяются 674 большие калории тепла.
Энергетический процесс дыхания у микробов протекает значительно сложнее и зависит от характера используемого питательного материала.
По типу дыхания микробов делят на аэробов и анаэробов, имеются микробы и с переходным типом дыхания.
Аэробы могут жить и развиваться при свободном доступе кислорода воздуха. Необходимую энергию для жизненных процессов они получают путем, поглощения кислорода и окисления питательных материалов.
Анаэробы способны развиваться без доступа кислорода. Свободный кислород воздуха на этих микробов оказывает вредное, губительное влияние. Строгие (облигатные) анаэробы (столбнячная палочка, возбудитель маслянокислого брожения) совсем не переносят кислорода. Необходимую энергию они получают путем расщепления органических веществ углеводов, белков, жиров, органических кислот, спиртов.
Факультативные анаэробы используют те же вещества, но применительно к условиям своего существования могут изменять анаэробный тип своего дыхания на аэробный. Так, дрожжи верхового брожения "при ограниченном притоке воздуха разлагают сахар на спирт и углекислоту; при обильной аэрации у них возникает аэробное дыхание с полным окислением сахара до углекислоты и воды. Молочнокислые бактерии в анаэробных условиях превращают глюкозу в молочную кислоту, при этом энергии освобождается несколько меньше, чем у аэробов. При недостатке кислорода денитрифицирующие бактерии пользуются для окисления органических соединений кислородом нитратов.
Приведенные примеры показывают разнообразие источников энергии и способов ее получения разными видами микробов; в силу этих причин у бактерий нет и быть но может единого механизма дыхания.
Большинство аэробных микроорганизмов окисляет органические питательные вещества в процессе дыхания до С02 и воды. Поскольку в молекуле СО 2 достигается высшая степень окисления углерода, в этом случае говорят о полном окислении и отличают этот тип дыхания от не полных окислений, при которых в качестве продуктов обмена выделяются частично окисленные органические соединения.
Под «полным окислением» имеется в виду лишь то, что не происходит вы деления каких-либо органических веществ; но это вовсе не означает, что окисляется весь поглощенный субстрат. В каждом случае значительная часть субстрата (40-70%) ассимилируется, т.е. превращается в вещества клеток.
Конечными продуктами «неполных окислений» могут быть уксусная, глюконовая, фумаровая, лимонная, молочная кислоты и ряд других соединений. Поскольку эти продукты сходны с теми, которые образуются при брожениях (пропионовая, масляная, янтарная, молочная кислоты и др.), а также в связи с тем, что при промышленных процессах брожения необходимы специальные технические устройства (ферментеры), неполные окисления называют также «окислительным брожением» или «аэробной ферментацией». Слова «брожение» и «ферментация» в этом случае отражают скорее технологический аспект.
Мы будем причислять к «неполным окислениям» также простое отщепление водорода от субстрата и использование микроорганизмов для катализа некоторых реакций, не имеющих для них какого-либо значения в обычных условиях. Ниже будут рассмотрены некоторые примеры таких окислений.
Дыхание бактерий
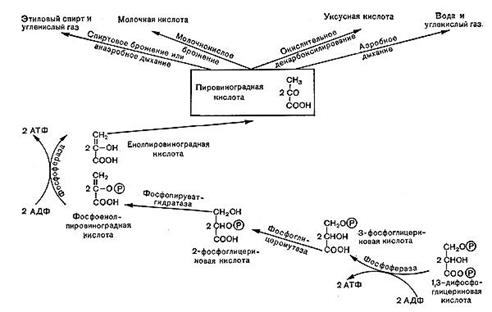
Рис.10. Роль пировнноградной кислоты в процессах дыхания и брожения.
Процесс дыхания заключается в том, что углеводы (или белки, жиры и другие запасные вещества клетки) разлагаются, окисляясь кислородом воздуха, до углекислого газа и воды. Выделяющаяся при этом энергия расходуется на поддержание жизнедеятельности организмов, рост и размножение. Бактерии вследствие ничтожно малых размеров своего тела не могут накапливать значительного количества запасных веществ. Поэтому они используют в основном питательные соединения среды.
В общем виде дыхание можно представить следующим уравнением:
С6Н12О6 + 602 = 6С02 + 6Н20 + 2,87-106 дж.
глюкоза кислород углекислый газ вода энергия
За этой простой формулой скрывается сложная цепь химических реакций, каждая из которых катализируется специфическим ферментом.
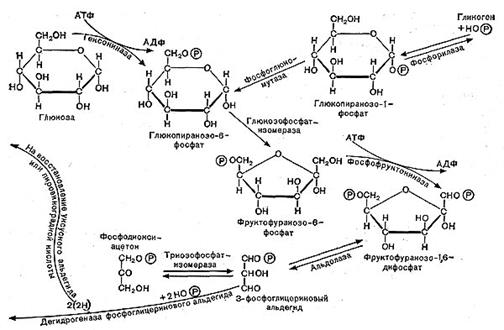
Рис. 11. Схема гликолитического пути расщепления углеводов.
Ферментативные реакции, происходящие в процессе дыхания, в настоящее время хорошо изучены. Схема реакций оказалась универсальной, т. е. в принципе одинаковой у животных, растений и многих микроорганизмов, в том числе бактерий. Процесс дыхания при окислении глюкозы складывается из следующих основных этапов (рис. 10).
Сначала происходит образование фосфорных эфиров глюкозы — моноaктивированная глюкоза в форме дифосфата далее расщепляется на два триозофосфата (трехуглеродные соединения): фосфоглицериновый альдегид и диоксиацетон-фосфат, которые могут обратимо превращаться друг в друга.
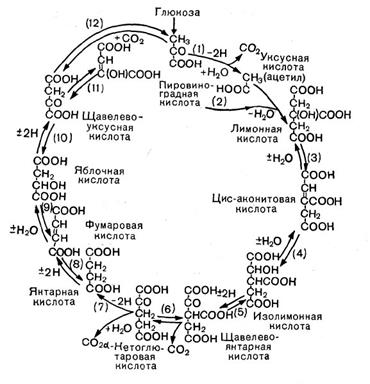
Рис. 12. Цикл трикарбоновых кислот. Стрелками показано направление, а номерами — порядок реакций.
Далее в обмен вступает фосфоглицериновый альдегид, он окисляется в дифосфо-глицериновую кислоту. Назначение этого процесса заключается в отщеплении атомов водорода от окисляемого субстрата и переносе водорода с помощью специфических окислительных ферментов к кислороду воздуха (см. рис. 10, 11).
Водород от фосфоглицеринового альдегида присоединяется к ферменту — никотин-амиддинуклеотиду (НАД); при этом альдегид окисляется до кислоты и выделяется энергия. Часть этой энергии тратится на образование АТФ; при этом присоединяется фосфорная кислота к аденозиндифосфат у— АДФ. При гидролизе АТФ энергия освобождается и может быть затрачена на различные процессы синтеза белка и другие нужды клетки.
Фосфоглицериыовая кислота окисляется до пировиноградной кислоты. При этом также образуется АТФ, т. е. запасается энергия.
На этом завершается первая — анаэробная — стадия процесса дыхания, которая носит название гликолитического пути или пути Эмбдена — Мейергофа — Парнаса. Для осуществления этих реакций кислород не требуется. Образовавшаяся пировиноградная кислота (СН3СОСООН) является интереснейшим и очень важным соединением. Пути расщепления глюкозы в процессе дыхания и многих брожений, вплоть до образования пировиноградной кислоты, идут совершенно одинаково, что впервые было установлено русским биохимиком С. П. Костычевым. Пировиноградная кислота является тем центральным пунктом, от которого расходятся пути дыхания и брожений, откуда начинается специфическая для данного процесса цепь ферментативных превращений -специфическая цепь химических реакций (рис. 11).
В процессе дыхания пировиноградная кислота вступает в цикл трикарбоновых кислот (рис. 12). Это сложный замкнутый круг превращений, в результате которых образуются органические кислоты с 4, 5 и 6 атомами углерода (яблочная, молочная, фумаро-вая, а-кетоглутаровая и лимонная) и отщепля-«тся углекислота.
Прежде всего от пировиноградной кислоты, содержащей три атома углерода, отщепляется СО2 — образуется уксусная кислота, которая с коферментом А образует активное соединение — ацетилкоэнзим А. Он передает остаток уксусной кислоты (ацетил) на щавелевоуксусную кислоту {4 атома углерода), и образуется лимонная кислота (6 атомов углерода). Лимонная кислота претерпевает несколько превращений, в результате выделяется С02 и образуется пяти-углеродное соединение — а-кетоглутаровая кислота. От нее тоже отщепляется С02 (третья молекула углекислого газа), и образуется янтарная кислота (4 атома углерода), которая затем превращается в фумаровую, яблочную и, наконец, щавелевоуксусную кислоту.
На этом цикл замыкается. Щавелевоуксусная кислота снова может вступить в цикл.
Таким образом, в цикл вступает трехуглерод-ная пировиноградная кислота, и по ходу превращений выделяются 3 молекулы С02.
Водород пировиноградной кислоты, освобождающийся при дегидрировании в аэробных условиях, не остается свободным — он поступает в дыхательную цепь (так же, как водород глицеринового альдегида, отнятый при превращении его в глицериновую кислоту). Это — цепь окислительных ферментов.
Ферменты, которые первыми берут на себя водород от окисляемого субстрата, называются первичными дегидрогеназами.
В их состав входят ди- или трипиридин-нуклео-тиды: НАД или НАДФ и специфический белок. Механизм присоединения водорода — один и тот же:
Окисляемое вещество — Н2 + НАД -> окисленное вещество + НАД*Н2
Водород, полученный дегидрогеназой, затем присоединяется к следующей ферментной системе — флавиновым ферментам (ФМН или ФАД).
От флавиновых ферментов электроны попадают на цитохромы — железосодержащие протеиды (сложные белки). По цепи цитохро-мов передается не атом водорода, а только электроны. При этом происходит изменение валентности железа: Fe++ — e->Fe++
Заключительная реакция дыхания — это присоединение протона и электрона к кислороду воздуха и образование воды. Но прежде происходит активирование молекулы кислорода под действием фермента цитохромоксидазы. Активирование сводится к тому, что кислород приобретает отрицательный заряд за счет присоединения электрона окисляемого вещества. К активированному кислороду присоединяется водород (протон), образуя воду.
Кроме упомянутой цепи переносчиков электронов и водорода, известны и другие. Процесс этот гораздо более сложен, чем изложенная схема.
Биологический смысл этих превращений заключается в окислении веществ и образовании энергии, В результате окисления молекулы сахара (глюкозы) в АТФ запасается 12,6-1053ж энергии, в самой молекуле сахара содержится 28,6-106 дж, следовательно, полезно используется 44% энергии. Это очень высокий коэффициент полезного действия, если сравнить его с к. п. д. современных машин.
В процессе дыхания образуется огромное количество энергии. Если вся она выделилась бы сразу, то клетка перестала бы существовать. Но этого не происходит, потому что энергия выделяется не вся сразу, а ступенчато, небольшими порциями. Выделение энергии небольшими дозами обусловлено тем, что дыхание представляет собой многоступенчатый процесс, на отдельных этапах которого образуются различные промежуточные продукты (с разной длиной углеродной цепочки) и выделяется энергия. Выделяющаяся энергия не расходуется в виде тепла, а запасается в универсальном макроэр-гическом соединении — АТФ. При расщеплении АТФ энергия может использоваться в любых процессах, необходимых для поддержания жизнедеятельности организма: на синтез различных органических веществ, механическую работу, поддержание осмотического давления протоплазмы и т. д.
Дыхание является процессом, дающим энергию, однако его биологическое значение этим не ограничивается. В результате химических реакций, сопровождающих дыхание, образуется большое количество промежуточных соединений. Из этих соединений, имеющих различное количество углеродных атомов, могут синтезироваться самые разнообразные вещества клетки: аминокислоты, жирные кислоты, жиры, белки, витамины.
Поэтому обмен углеводов определяет остальные обмены веществ (белков, жиров). В этом его огромное значение.
С процессом дыхания, его химическими реакциями связано одно из удивительных свойств микробов — способность испускать видимый свет — люминесцировать.
Известно, что ряд живых организмов, в том числе бактерии, могут испускать видимый свет. Люминесценция, вызываемая микроорганизмами, известна уже в течение столетий. Скопление люминесцирующих бактерий, находящихся в симбиозе с мелкими морскими животными, иногда приводит к свечению моря; с люминесценцией встречались также при росте некоторых бактерий на мясе и т. д.
К основным компонентам, взаимодействие между которыми приводит к испусканию света, относятся восстановленные формы ФМН или НАД, молекулярный кислород, фермент люцифераза и окисляемое соединение — люциферин. Предполагается, что восстановленные НАД или ФМН реагируют с люциферазой, кислородом и люциферином, в результате чего электроны в некоторых молекулах переходят в возбужденное состояние и возвращение этих электронов на основной уровень сопровождается испусканием света. Люминесценцию у микробов рассматривают как «расточительный процесс», так как при этом энергетическая эффективность дыхания снижается.
Поможем написать любую работу на аналогичную тему