Рецептором называют специализированную клетку, эволюционно приспособленную к восприятию из внешней или внутренней среды определенного раздражителя и к преобразованию его энергии из физической или химической формы в форму нервного возбуждения.
КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ
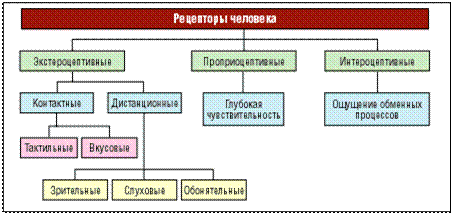 Классификация рецепторов основывается, в первую очередь, на характере ощущений, возникающих у человека при их раздражении. Различают зрительные, слуховые, обонятельные, вкусовые, осязательные рецепторы, терморецепторы, проприои вестибулорецепторы (рецепторы положения тела и его частей в пространстве). Обсуждается вопрос существования специальных рецепторов боли.
Классификация рецепторов основывается, в первую очередь, на характере ощущений, возникающих у человека при их раздражении. Различают зрительные, слуховые, обонятельные, вкусовые, осязательные рецепторы, терморецепторы, проприои вестибулорецепторы (рецепторы положения тела и его частей в пространстве). Обсуждается вопрос существования специальных рецепторов боли.
Рецепторы по месту расположения разделяют на внешние, или экстерорецепторы, и внутренние, или интерорецепторы. К экстерорецепторам относятся слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам относятся вестибулорецепторы и проприорецепторы (рецепторы опорно-двигательного аппарата), а также интерорецепторы, сигнализирующие о состоянии внутренних органов.
По характеру контакта с внешней средой рецепторы делятся на дистантные, получающие информацию на расстоянии от источника раздражения (зрительные, слуховые и обонятельные), и контактные – возбуждающиеся при непосредственном соприкосновении с раздражителем (вкусовые и тактильные).
В зависимости от природы вида воспринимаемого раздражителя, на который они оптимально настроены, различают пять типов рецепторов.
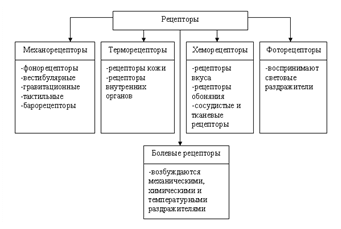 Механорецепторы возбуждаются при их механической деформации; расположены в коже, сосудах, внутренних органах, опорно-двигательном аппарате, слуховой и вестибулярной системах.
Механорецепторы возбуждаются при их механической деформации; расположены в коже, сосудах, внутренних органах, опорно-двигательном аппарате, слуховой и вестибулярной системах.- Хеморецепторы воспринимают химические изменения внешней и внутренней среды организма. К ним относятся вкусовые и обонятельные рецепторы, а также рецепторы, реагирующие на изменение состава крови, лимфы, межклеточной и цереброспинальной жидкости (изменение напряжения О2 и СО2, осмолярности и рН, уровня глюкозы и других веществ). Такие рецепторы есть в слизистой оболочке языка и носа, каротидном и аортальном тельцах, гипоталамусе и продолговатом мозге.
- Терморецепторы реагируют на изменения температуры. Они подразделяются на тепловые и холодовые рецепторы и находятся в коже, слизистых оболочках, сосудах, внутренних органах, гипоталамусе, среднем, продолговатом и спинном мозге.
- Фоторецепторы в сетчатке глаза воспринимают световую (электромагнитную) энергию.
- Ноцицепторы, возбуждение которых сопровождается болевыми ощущениями (болевые рецепторы). Раздражителями этих рецепторов являются механические, термические и химические (ги-стамин, брадикинин, К+, Н+ и др.) факторы. Болевые стимулы воспринимаются свободными нервными окончаниями, которые имеются в коже, мышцах, внутренних органах, дентине, сосудах. С психофизиологической точки зрения рецепторы подразделяют в соответствии с органами чувств и формируемыми ощущениями на зрительные, слуховые, вкусовые, обонятельные и тактильные.
В зависимости от строения рецепторов их подразделяют на первичные, или первичночувствующие, которые являются специализированными окончаниями чувствительного нейрона, и вторичные, или вторичночувствующие, представляющие собой клетки эпителиального происхождения, способные к образованию рецепторного потенциала в ответ на действие адекватного стимула.
Первичночувствующие рецепторы могут сами генерировать потенциалы действия в ответ на раздражение адекватным стимулом, если величина их рецепторного потенциала достигнет пороговой величины. К ним относятся обонятельные рецепторы, большинство механорецепторов кожи, терморецепторы, болевые рецепторы или ноцицепторы, проприоцепторы и большинство интерорецепторов внутренних органов. Тело нейрона расположено в спинно-мозговом ганглии или в ганглии черепных нервов. В первичном рецепторе раздражитель действует непосредственно на окончания сенсорного нейрона. Первичные рецепторы являются филогенетически более древними структурами, к ним относятся обонятельные, тактильные, температурные, болевые рецепторы и проприорецепторы.
Вторичночувствующие рецепторы отвечают на действие раздражителя лишь возникновением рецепторного потенциала, от величины которого зависит количество выделяемого этими клетками медиатора. С его помощью вторичные рецепторы действуют на нервные окончания чувствительных нейронов, генерирующих потенциалы действия в зависимости от количества медиатора, выделившегося из вторичночувствующих рецепторов. Во вторичных рецепторах имеется специальная клетка, синаптически связанная с окончанием дендрита сенсорного нейрона. Это клетка, например фоторецептор, эпителиальной природы или нейроэктодермального происхождения. Вторичные рецепторы представлены вкусовыми, слуховыми и вестибулярными рецепторами, а также хемочувствительными клетками синокаротидного клубочка. Фоторецепторы сетчатки, имеющие общее происхождение с нервными клетками, чаще относят к первичным рецепторам, но отсутствие у них способности генерировать потенциалы действия указывает на их сходство с вторичными рецепторами.
По скорости адаптации рецепторы делят на три группы: быстро адаптирующиеся (фазные), медленно адаптирующиеся (тонические) и смешанные (фазнотонические), адаптирующиеся со средней скоростью. Примером быстро адаптирующихся рецепторов являются рецепторы вибрации (тельца Пачини) и прикосновения (тельца Мейснера) к коже. К медленно адаптирующимся рецепторам относятся проприорецепторы, рецепторы растяжения легких, болевые рецепторы. Со средней скоростью адаптируются фоторецепторы сетчатки, терморецепторы кожи.
Большинство рецепторов возбуждаются в ответ на действие стимулов только одной физической природы и поэтому относятся к мономодальным. Их можно возбудить и некоторыми неадекватными раздражителями, например фоторецепторы — сильным давлением на глазное яблоко, а вкусовые рецепторы — прикосновением языка к контактам гальванической батареи, но получить качественно различаемые ощущения в таких случаях невозможно.
Наряду с мономодальными существуют полимодальные рецепторы, адекватными стимулами которых могут служить раздражители разной природы. К такому типу рецепторов принадлежат некоторые болевые рецепторы, или ноцицепторы (лат. nocens — вредный), которые можно возбудить механическими, термическими и химическими стимулами. Полимодальность имеется у терморецепторов, реагирующих на повышение концентрации калия во внеклеточном пространстве так же, как на повышение температуры.
ОБЩИЕ МЕХАНИЗМЫ ВОЗБУЖДЕНИЯ РЕЦЕПТОРОВ
При действии стимула в рецепторе происходит преобразование энергии внешнего раздражения в рецепторный сигнал (трансдукция сигнала). Этот процесс включает в себя три основных этапа:
1) взаимодействие стимула с рецепторной белковой молекулой, которая находится в мембране рецептора;
2) усиление и передачу стимула в пределах рецепторной клетки
3) открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток, что, как правило, приводит к деполяризации клеточной мембраны рецепторной клетки (возникновению так называемого рецепторного потенциала).
Чувствительность рецепторных элементов к адекватным раздражителям, к восприятию которых они эволюционно приспособлены, предельно высока. Так, обонятельный рецептор может возбудиться при действии одиночной молекулы пахучего вещества, фоторецептор – при действии одиночного кванта света.
Механизм возбуждения рецепторов связан с изменением проницаемости клеточной мембраны для ионов калия и натрия. Когда раздражение достигает пороговой величины, возбуждается сенсорный нейрон, посылающий импульс в центральную нервную систему. Можно сказать, что рецепторы кодируют поступающую информацию в виде электрических сигналов. Сенсорная клетка посылает информацию по принципу «всё или ничего» (есть сигнал / нет сигнала). При действии стимула на рецепторную клетку в белково-липидном слое мембраны происходит изменение пространственной конфигурации белковых рецепторных молекул. Это приводит к изменению проницаемости мембраны для определенных ионов, чаще всего для ионов натрия, но в последние годы открыта еще и роль калия в этом процессе. Возникают ионные токи, изменяется заряд мембраны и происходит генерация рецепторного потенциала (РП). А далее процесс возбуждения протекает в разных рецепторах по-разному.
В первично чувствующих рецепторах, которые являются свободными голыми окончаниями чувствительного нейрона (обонятельных, тактильных, проприоцептивных), РП воздействует на соседние, наиболее чувствительные участки мембраны, где генерируется потенциал действия (ПД), который далее в виде импульсов распространяется по нервному волокну. Таким образом, когда рецепторный потенциал достигает определенной величины, на его фоне возникает распространяющийся ПД. Преобразование энергии внешнего стимула в ПД в первичных рецепторах может происходить как непосредственно на мембране, так и при участии некоторых вспомогательных структур.
Рецепторный и распространяющийся потенциалы возникают в первичных рецепторах в одних и тех же элементах. Так, в расположенных в коже окончаниях отростка сенсорного нейрона при действии раздражителя сначала формируется рецепторный потенциал, под влиянием которого в ближайшем перехвате Ранвье возникает распространяющийся потенциал. Следовательно, в первичных рецепторах рецепторный потенциал является причиной возникновения — генерации — распространяющегося ПД, поэтому его называют еще генераторным
Так, например, происходит в тельце Пачини. Рецептор здесь представлен голым окончанием аксона, которое окружено соединительнотканной капсулой. При сдавливании тельца Пачини регистрируется РП, который далее преобразуется в импульсный ответ афферентного волокна.
Во вторично чувствующих рецепторах, которые представлены специализированными клетками (зрительные, слуховые, вкусовые, вестибулярные), РП приводит к образованию и выделению медиатора из пресинаптического отдела рецепторной клетки в синаптическую щель рецепторно-афферентного синапса. Этот медиатор воздействует на постсинаптическую мембрану чувствительного нейрона, вызывает ее деполяризацию и образование постсинаптического потенциала, который называют генераторным потенциалом (ГП). ГП, воздействуя на внесинаптические участки мембраны чувствительного нейрона, обусловливает генерацию ПД. ГП может быть как де-, так и гиперполяризационным и соответственно вызывать возбуждение или тормозить импульсный ответ афферентного волокна.
Во вторичных рецепторах рецепторный и генераторный потенциалы не являются однозначными понятиями: рецепторный потенциал возникает в рецепторных образованиях, а генераторный—в конечных разветвлениях отростка сенсорного нейрона. Наличие генераторных потенциалов было установлено для всех рецепторных образований. Это позволило сделать вывод о том, что пусковым механизмом для возникновения нервных импульсов является генераторный потенциал рецепторов. Его амплитуда возрастает в логарифмической зависимости от интенсивности раздражена.
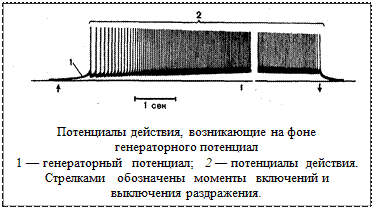 В свою очередь, частота возникающих распространяющихся импульсов прямо пропорциональна величине генераторного потенциала, так что в конечном результате она находится тоже в логарифмической зависимости от интенсивности раздражения. Следовательно, уже на периферии происходит анализ раздражителя по силе: чем сильнее раздражитель, тем больше генераторный потенциал и частота возникающих потенциалов действия и тем интенсивнее возникающее ощущение.
В свою очередь, частота возникающих распространяющихся импульсов прямо пропорциональна величине генераторного потенциала, так что в конечном результате она находится тоже в логарифмической зависимости от интенсивности раздражения. Следовательно, уже на периферии происходит анализ раздражителя по силе: чем сильнее раздражитель, тем больше генераторный потенциал и частота возникающих потенциалов действия и тем интенсивнее возникающее ощущение.
Спонтанная активность рецепторных образований и их реакции на раздражение. Для всех рецепторных образований характерна спонтанная, или самопроизвольная, активность. Она проявляется в генерации ПД без нанесения раздражений. Спонтанная активность служит источником деятельного состояния ретикулярной формации и создает возможность проявлять реакции на раздражения в двух направлениях: возбуждения и торможения. При возбуждении активность усиливается, при торможении уменьшается. Это обеспечивает известную гибкость системы, воспринимающей различные раздражения.
Торможение и адаптация. Для деятельности анализаторов характерно наличие торможения уже в периферическом его отделе. Торможение обнаружено в рецепторных образованиях разных анализаторов и у разных представителей мира животных. Торможение в рецепторных образованиях органов чувств способствует периферическому анализу раздражений. Так, в зрительном анализаторе оно обеспечивает контрастность изображения путем подчеркивания линий и контуров предметов.
Всем рецепторам свойственна адаптация к действующему раздражителю. Адаптацией называют снижение чувствительности анализатора к постоянно действующему раздражению. Субъективно она проявляется в уменьшении интенсивного ощущения или в полном его исчезновении, а объективно — в уменьшении числа импульсов, идущих по афферентному нерву раздражаемого анализатора. Раздражитель, действующий постоянно, перестает раздражать. Поэтому в помещении, где имеется какое-либо пахучее вещество, сначала очень сильно ощущение его запаха, а через некоторое время оно исчезает. Биологическое значение адаптации заключается в том, что нейроны центральной нервной системы, быстро освобождаясь от одного вида деятельности, способны все время воспринимать новые раздражения, т. е. осуществлять реакцию на изменения, происходящие в окружающей среде.
 В разных рецепторах скорость адаптации неодинакова. Очень быстро возникает адаптация в рецепторных образованиях глаза и уха, а в двигательном анализаторе ее скорость невелика, так как постоянная импульсация от рецепторов мышц необходима для коррекции движения.
В разных рецепторах скорость адаптации неодинакова. Очень быстро возникает адаптация в рецепторных образованиях глаза и уха, а в двигательном анализаторе ее скорость невелика, так как постоянная импульсация от рецепторов мышц необходима для коррекции движения.
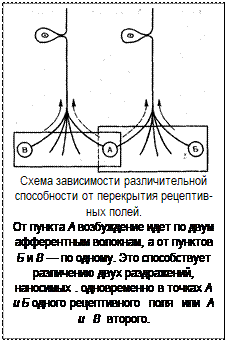
Рецептивное поле анализатора и чувствительность рецепторных образований. Для каждого анализатора характерно наличие цептивного поля. Им называют воспринимающий раздражения участок поверхности, в котором разветвляется афферентное нервное волокно одной нервной клетки. Площадь рецептивных участков различна не только для разных анализаторов, но широко варьирует в пределах каждого из них.
С величиной рецептивного поля связана чувствительность действующему раздражителю. Экспериментально было показано, что для возникновения возбуждения в нейроне недостаточно активности одного рецепторного элемента. При действии раздражителя происходит суммация эффектов раздражения в элементах данного рецептивного поля.
Причем установлено, что интенсивность раздражителя в известных пределах взаимозаменяема с площадью раздражаемого участка. Чем больше площадь суммации, тем менее интенсивным может быть раздражитель для вызова одного и того же эффекта. Таким образом, величиной рецептивного поля определяется интенсивность возникающей реакции и способность к восприятию слабых раздражений.
Способность различать две точки пространства как раздельные при увеличении рецептивного поля, наоборот, падает. При большом рецептивном поле одновременно нанесение раздражения на две его точки, даже далеко расположенные, воспринимается как одно, ибо они адресуются к одной и той же нервной клетке. Недостаточная различительная способность частично компенсируется взаимным перекрытием рецептивных полей.
Чувствительность рецепторных образований. Все рецепторные образования очень чувствительны к адекватным раздражениям, и нечувствительны к неадекватным.
Поможем написать любую работу на аналогичную тему
Реферат
Рецепторы и их классификация. Закономерности деятельности рецепторных образований.
От 250 руб
Контрольная работа
Рецепторы и их классификация. Закономерности деятельности рецепторных образований.
От 250 руб
Курсовая работа
Рецепторы и их классификация. Закономерности деятельности рецепторных образований.
От 700 руб