Проприоцепция - это группа сигналов, посылаемых в центральную нервную систему специальными терминалами (проприоцепторами), расположенными в суставных капсулах, связках, сухожилиях и мышцах.
Проприоцепторы, среди которых выделяют мышечные рецепторы, или мышечные веретена (рецепторы растяжения), сухожильные рецепторы, или органы Гольджи (рецепторы мышечной силы), а также суставные рецепторы относятся к механорецепторам, посылающим в ЦНС информацию о положении, деформации и смещениях различных частей тела.
Функционирование этих рецепторов обеспечивает координацию всех подвижных органов и тканей животного и человека в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов животные теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
Если человек закроет глаза и попытается написать текст, то буквы будут написаны достаточно четко. Этим простым способом легко убедиться в умении человека пользоваться информацией, идущей от мышц и суставов.
Проприоцепторы составляют периферическую часть проприоцептивной сенсорной системы, или двигательного анализатора. Вместе с тем, несмотря на то, что миллионы людей ежедневно пользуются услугами этого анализатора, мы до сегодняшнего дня знаем сравнительно мало о его деятельности. Это особенно касается работы коркового отдела двигательного анализатора. Внутренние проприоцепторы находятся в мышцах, сухожилиях, сухожильных влагалищах, межкостных мембранах, фасциях, тканях суставов, надкостнице и т.д. Среди них имеются неспециализированные рецепторы, встречающиеся и в других частях тела (свободные нервные окончания, инкапсулированные рецепторы типа телец Руффини и Пачини), и специализированные — мышечные веретена и сухожильные органы (или рецепторы) Гольджи.
Двигательный, или кинестетический, анализатор (мышечная сенсорная система) обеспечивает формирование так называемого мышечного чувства при изменении напряжения мышц, суставных сумок, связок и сухожилий. Проблема мышечно-суставных ощущений имеет исключительное значение для физиологии и психологии. Специфические особенности человека появились благодаря более совершенной организации мышечной сенсорной системы по сравнению с животными.
И.М. Сеченов считал, что мышечное чувство является ближайшим регулятором движений и одним из орудий ориентации человека в пространстве и времени. Ощущения положения и перемещения тела в пространстве, ощущения во время трудовой деятельности и членораздельная речь лежат в основе формирования сознания человека и его представлений об окружающем мире.
Мышечное чувство обладает тремя качествами. Это, во-первых, ощущение положения конечностей, когда человек может определить положение своих конечностей и их частей относительно друг друга. Во-вторых, ощущение движения, когда, изменяя угол сгибания в суставе, человек осознаёт скорость и направление движения. Третьим качеством является ощущение усилия, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определённом положении при подъёме или перемещении груза.
Наряду с кожной, зрительной и вестибулярной сенсорными системами двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности.
Двигательная сенсорная система служит для анализа состояния двигательного аппарата — его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз.
Общий план организации. Двигательная сенсорная система состоит из следующих 3-х отделов:
- периферический отдел, представленный проприорецепторами, расположенными в мышцах, сухожилиях и суставных сумках;
- проводниковый и отдел, который начинается биполярными клетками (первыми нейронами), тела которых расположены вне ЦНС — в спинномозговых узлах. Один их отросток связан с рецепторами, другой входит в спинной мозги передает проприоцептивные импульсы ко вторым нейронам в продолговатый мозг(часть путей от проприорецепторов направляется в кору мозжечка), а далее к третьим нейронам — релейным ядрам таламуса (в промежуточный мозг);
- корковый отдел находится в передней центральной извилине коры больших полушарий.
Функции проприорецепторов
К проприорецепторам относятся мышечные веретена, сухожильные органы (или органы Гольджи) и суставные рецепторы (рецепторы суставной капсулы и суставных связок). Все эти рецепторы представляют собой механорецепторы, специфическим раздражителем которых является их растяжение.
Мышечные веретена человека, представляют собой продолговатые образования длиной несколько миллиметров, шириной десятые доли миллиметра, которые расположены в толще мышцы. В разных скелетных мышцах число веретен на 1 г ткани варьирует от нескольких единиц до сотни.
Таким образом, мышечные веретена как датчики состояния силы мышцы и скорости ее растяжения реагируют на два воздействия: периферическое — изменение длины мышцы, и центральное — изменение уровня активации гамма-мотонейронов. Поэтому реакции веретен в условиях естественной деятельности мышц довольно сложны. При растяжении пассивной мышцы наблюдается активация рецепторов веретен; она вызывает миотатичес-кий рефлекс, или рефлекс на растяжение. При активном сокращении мышцы уменьшение ее длины оказывает на рецепторы веретена дезактивирующее действие, а возбуждение гамма-мотонейронов, сопутствующее возбуждению альфа-мотонейронов, приводит к реактивации рецепторов. Вследствие этого импульсация от рецепторов веретен во время движения зависит от длины мышцы, скорости ее укорочения и силы сокращения.
Сухожильные органы (рецепторы) Гольджи человека располагаются в зоне соединения мышечных волокон с сухожилием, последовательно по отношению к мышечным волокнам.
Сухожильные органы представляют собой структуру вытянутой веретенообразной или цилиндрической формы, длина которой у человека может достигать 1 мм. Этот первичночувствующий рецептор. В условиях покоя, т.е. когда мышца не сокращена, от сухожильного органа идет фоновая импульсация. В условиях мышечного сокращения частота импульсации возрастает прямо пропорционально величине мышечного сокращения, что позволяет рассматривать сухожильный орган как источник информации о силе, развиваемой мышцей. В тоже время сухожильный орган слабо реагирует на растяжение мышцы.
В результате последовательного крепления сухожильных органов к мышечным волокнам (а в ряде случаев — к мышечным веретенам), растяжение сухожильных механорецепторов происходит при напряжении мышц. Таким образом, в отличие от мышечных веретен, сухожильные рецепторы информируют нервные центры о степени напряжения мыши, и скорости его развития.
Суставные рецепторы реагируют на положение сустава и на изменения суставного угла, участвуя, таким образом, в системе обратных связей от двигательного аппарата и в управлении им. Суставные рецепторы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецепторы посылают информацию о величине суставного угла, т. е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецепторы возбуждаются только в момент движения в суставе, т. е. посылают информацию о скорости движения. Частота их импульсации возрастаете увеличением скорости изменения суставного угла.
 Проводниковый и корковый отделы проприоцеп-тивного анализатора млекопитающих и человека. Информация от мышечных, сухожильных и суставных рецепторов поступает через аксоны первых афферентных нейронов, находящихся в спинномозговых ганглиях, в спинной мозг, где частично переключается на альфа-мотонейроны или на вставочные нейроны (например, на клетки Реншоу), а частично направляется по восходящим путям в высшие отделы головного мозга. В частности, по путям Флексига и Говерса проприоцептивная импульсация доставляется к мозжечку, а по пучкам Голля и Бурдаха, проходящим в дорсальных канатиках спинного мозга, она доходит до нейронов одноименных ядер, расположенных в продолговатом мозге.
Проводниковый и корковый отделы проприоцеп-тивного анализатора млекопитающих и человека. Информация от мышечных, сухожильных и суставных рецепторов поступает через аксоны первых афферентных нейронов, находящихся в спинномозговых ганглиях, в спинной мозг, где частично переключается на альфа-мотонейроны или на вставочные нейроны (например, на клетки Реншоу), а частично направляется по восходящим путям в высшие отделы головного мозга. В частности, по путям Флексига и Говерса проприоцептивная импульсация доставляется к мозжечку, а по пучкам Голля и Бурдаха, проходящим в дорсальных канатиках спинного мозга, она доходит до нейронов одноименных ядер, расположенных в продолговатом мозге.
Аксоны таламических нейронов (нейронов третьего порядка) оканчиваются в коре больших полушарий, главным образом, в соматосенсорной коре (постцентральная извилина) и в области сильвие-вой борозды (соответственно, участки S-1 и S-2), а также частично в двигательной (префронтальной) области коры. Эта информация используется двигательными системами мозга достаточно широко, в том числе для принятия решения о замысле движения, а также для его реализации. Кроме того, у человека на основе про-приоцептивной информации формируются представления о состоянии мышц и суставов, а также, в целом, о положении тела в пространстве.
Сигналы, идущие от рецепторов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецепторов кожи, называют кинестетическими, т. е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецепторов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприорецепторов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз.
Таким, образом, проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза.
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов.
Вестибулярный анализатор - анализатор, обеспечивающий анализ информации о положении и перемещениях тела в пространстве.
лимбическая система
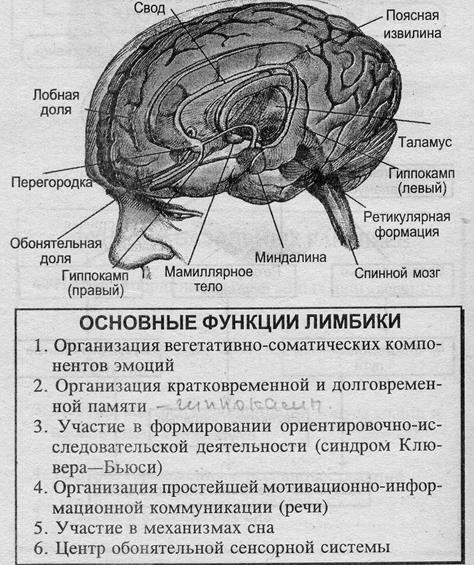
Лимбические круги
Большой круг Пейпса:
Гиппокамп → свод → мамиллярные тела → мамиллярно-таламический пучок Вик-д,Азира → таламус → поясная извилина → гиппокамп.
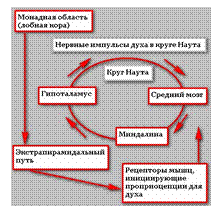
Малый круг Наута:
Миндалина → конечная полоска → гипоталамус → перегородка → миндалина.
Функциональная структура лимбики по МакЛИНУ (1970)
- Нижний отдел – миндалина и гиппокамп – центры эмоций и поведения для выживания и самосохранения
- Верхний отдел – поясная извилина и височная кора – центры общительности и сексуальности
- Средний отдел – гипоталамус и поясная извилина – центры биосоциальных инстинктов
Эмоциональный мозг
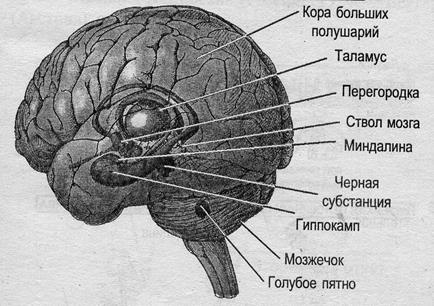
Эффекты поражения стриопаллидарной системы
- Поражения хвостатого ядра – гиперкинезы-атетозы и хорея (пляска святого Витта)
- Поражения паллидум – обеднение двигательной активности при повышенном пластическом тонусе и треморе – Болезнь Паркинсона
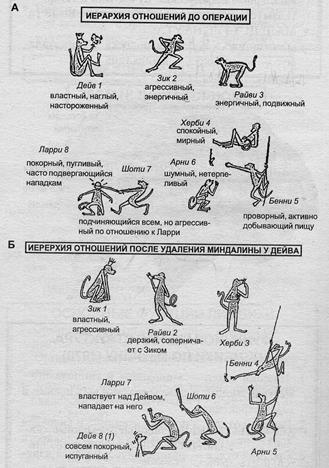
Роль миндалины в иерархическом поведении
(по К.Прибрам)
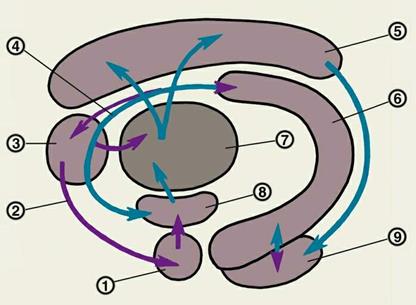
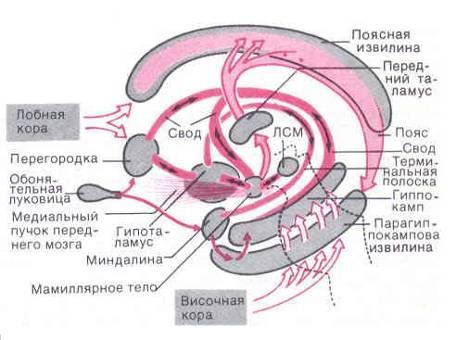
Морфофункциональная характеристика лимбической системы — схема взаимодействия структур круга Пейпса: 1 — амигдалоидная область; 2 — обонятельная система; 3 — перегородка; 4 — свод 5 — поясная извилина 6 — гиппокамп 7 — переднее ядро таламуса 8 — гипоталамус 9 — энторинальная кора; синими стрелками обозначены морфологические связи круга Пейпса, фиолетовыми — связи, не входящие в него.
Круги Пейпса и Наута
Функции лимбической системы мозга, состоящей из кругов Пейпса и Наута, в психических явлениях человека и высших животных и являются предметом познания психофизиологии. Психофизиология и нейропсихология - вот две базовые теории в построении общей психологии.
Важной структурой круга Пейпса является гиппокамп, благодаря которому поддерживается информация о текущих событиях. По орбитам круга Пейпса циркулируют паттерны - коды информации из нервных импульсов. Эти паттерны через таламус и поясную извилину передаются всей коре больших полушарий.
Круг Наута проходит через гипоталамус, через который психика управляет всей гормональной активностью организма. Круг Пейпса оперирует информацией, в круге Наута формируются мотивы ответного поведения. Оба круга взаимосвязаны через связь гипоталамуса с мамиллярным телом.
Круг Пейпса. Нейрофизиологам известна лимбическая система мозга (архипалеокортекс) под новой корой больших полушарий (неокортекс). Эту систему ещё называют эмоциональным мозгом - её мы и возьмём за основу для объяснения психических явлений человека.
Лимбическая система состоит из огромного множества замкнутых нервных волокон в которых циркулируют (реверберируют) нервные импульсы. Назовём одно такое волокно (замкнутую цепь нейронных аксонов) орбитой, систему нервных импульсов в орбите с определённым информационным кодом паттерном.
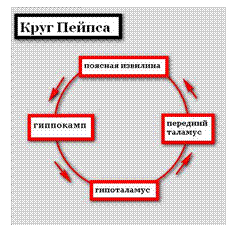 Круг Пейпса лимбической системы состоит из функционально важных для психики человека структур: гиппокамп, гипоталамус, передний таламус, поясная извилина.
Круг Пейпса лимбической системы состоит из функционально важных для психики человека структур: гиппокамп, гипоталамус, передний таламус, поясная извилина.
Поясная извилина имеет связь со всей корой больших полушарий. Гиппокамп является информационным центром лимбической системы . Гиппокамп имеет два вида нейронов, одни из которых поддерживают в памяти незнакомую информацию в течение 30 дней. Паттерн ы незнакомой информации от гиппокампа идут к гипоталамусу, повышая его гормональную активность и эмоциональное состояние, но в таламусе они прерываются. Таламус есть своего рода фильтр, который пропускает только известную информацию и отправляет её в поясную извилину, в неокортекс для формирования ответного поведения.
Не всегда следует с получением сведения сразу же запускать поведение, необходимо выждать время. И вся информация, отфильтрованная таламусом, реверберирует в круге Пейпса, её поддерживает второй вид нейронов гиппокампа. Паттерны, совершающие полный оборот по кругу Пейпса, есть сознание. Сознание есть вся совокупность оперативной информации, необходимая для регулирования текущего поведения. Как только поведение достигает определённой цели, имеющееся сознание в лимбической системе затухает, человек переходит к новому сознанию.
Гиппокамп, пропуская через себя сознание, одновременно поддерживает и подсознание, то есть те паттерны, которые не пропускает таламус, не укладываются они в его логических связях. Подсознание действует на гипоталамус, обуславливает состояние тревожности, но человек его не осмысливает, не понимает.
Вся совокупность сведений о предмете фиксируется в матрице. Матрица состоит из орбит и паттернов в этих орбитах. Причём, как и в матрице из линейной алгебры, на каждой орбите (строке) определённое количество паттернов (элементов). Достигается это в результате латерального торможения: если орбита более возбуждённая, имеет высокую плотность паттернов, то она доминирует и затормаживает соседние орбиты. Таким образом, в матрице сохраняются орбиты с равными плотностями паттернов. Менее возбуждённые орбиты отражают сведения о предмете, к которым субъективно мало интереса - и они выпадают из матрицы, из сознания.
Выпадение орбит из матрицы назовём редукцией. Редукция бывает эмоциональной и логической. Эмоциональную редукцию осуществляют доминирующие орбиты, затормаживающие орбиты с меньшим возбуждением. Логическая редукция протекает в структуре таламуса посредством нейронов с тормозным медиатором. Если два каких-то сведения исключают третье сведение, то соответствующие им орбиты затормаживают третью орбиту.
Вообще, ядра таламуса, имеющие связь с лимбической системой , назовём рассудком. Рассудок имеет наследственные или приобретённые логические связи из нейронов, которые обрабатывают сведения о предмете и направляют их к определённым нейронам неокортекса.
Функциональной единицей неокортекса является не нейрон, а колонка, состоящая из звёздчатых и одного пирамидного нейрона, расположенных друг над другом в шести слоях коры. У каждой матрицы есть определитель - это колонка неокортекса, соответствующая предмету.
 Матрица реверберирует по кругу Пейпца , находится в сознании и одновременно поддерживает активность определителя. Если матрица не поддерживает активность определителя, то определитель спит и никак не реагирует на предмет восприятия.
Матрица реверберирует по кругу Пейпца , находится в сознании и одновременно поддерживает активность определителя. Если матрица не поддерживает активность определителя, то определитель спит и никак не реагирует на предмет восприятия.
Круг Наута в лимбической системе имеет меньший размер , чем круг Пейпца , но функциональное значение его гораздо выше.
Реверберируют по кругу нервные импульсы проприоцепций из рецепторов мышц. Сокращает рецепторы мышц лобная кора неокортекса.
Если таламус есть рассудок, то лобная кора (монадная область) есть разум человека.
Классификация рецепторов.
В основу классификации рецепторов положено несколько критериев.
v Психофизиологический характер ощущения: тепловые, холодовые, болевые и др.
v Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осмбрецепторы и др.
v Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы.
v Отношение к одной или нескольким модальностям: моно- и полимодальные (мономодальные преобразуют в нервный импульс только один вид раздражителя - световой, температурный и т. д., полимодальные могут несколько раздражителей преобразовать в нервный импульс - механический и температурный, механический и химический и т. д.).
v Способность воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним: контактные и дистантные.
v Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы).
v Скорость адаптации: быстроадаптирующиеся, (тактильные), медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы).
v Отношение к различным моментам действия раздражителя: при включении раздражителя, при его выключении, на протяжении всего времени действия раздражителя.
v Морфофункциональная организация и механизм возникновения возбуждения: первичночувствующие и вторичночувствующие.
В первичночувствующих рецепторах стимул действует на воспринимающий субстрат, заложенный в самом сенсорном нейроне, который при этом возбуждается непосредственно (первично) раздражителем. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена.
К вторичночувствующим относятся те рецепторы, у которых между действующим стимулом и сенсорным нейроном располагаются дополнительные рецептирующие клетки, при этом сенсорный нейрон возбуждается не непосредственно стимулом, а опосредовано (вторично) - потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
Механизм возникновения возбуждения у этих рецепторов различен. В первичночувствующем рецепторе транформация энергии раздражителя и возникновение импульсной активности идет в самом сенсорном нейроне. У вторичночувствующих рецепторов между сенсорным нейроном и стимулом расположена рецептирующая клетка, в которой под влиянием раздражителя идут процессы трансформации энергии раздражителя в процесс возбуждения. Но в этой клетке не возникает импульсной активности. Рецепторные клетки синапсами соединены с сенсорными нейронами. Под влиянием потенциала рецептирирующей клетки выделяется медиатор, который возбуждает нервное окончание сенсорного нейрона и вызывает в нем появление локального ответа - постсинаптического потенциала. Он оказывает деполяризующее действие на отходящее нервное волокно, в котором возникает импульсная активность.
Следовательно, у вторичночувствующих рецепторов локальная деполяризация возникает дважды: в рецептирующей клетке и в сенсорном' нейроне. Поэтому принято называть градуальный электрический ответ рецептирующей клетки рецепторным потенциалом, а локальную деполяризацию сенсорного нейрона генераторным потенциалом, имея в виду, что он генерирует в отходящем от рецептора нервном волокне распространяющееся возбуждение. У первичночувствующих рецепторов рецепторный потенциал является и генераторным. Таким образом, рецепторный акт можно изобразить в виде следующей схемы.
Для первичночувствующих рецепторов:
I этап - специфическое взаимодействие раздражителя с мембраной рецептора;
II этап - возникновение рецепторного потенциала в месте взаимодействия раздражителя с рецептором в результате изменения проницаемости мембраны для ионов натрия (или кальция);
III этап - электротоническое распространение рецепторного потенциала к аксону сенсорного нейрона (пассивное распространение рецепторного потенциала вдоль нервного волокна называется электротоническим);
IV этап - генерация потенциала действия;
V этап - проведение потенциала действия по нервному волокну в ортодромном направлении.
Для вторичночувствующих рецепторов:
I-III этапы совпадают с такими же этапами первичночувствующих рецепторов, но протекают они в специализированной рецептирующей клетке и заканчиваются на ее пресинаптической мембране;
IV этап - выделение медиатора пресинаптическими структурами рецептирующей клетки;
V этап - возникновение генераторного потенциала на постсинаптической мембране нервного волокна;
VI этап - электротоническое распространение генераторного потенциала по нервному волокну;
VII этап - генерация потенциала действия электрогенными участками нервного волокна;
VIII этап - проведение потенциала действия по нервному волокну в ортодромном направлении.
Слуховые функции.
Анализ частоты звука (высоты тона). Звуковые колебания разной частоты вовлекают в колебательный процесс основную мембрану на всем ее протяжении неодинаково. Локализация амплитудного максимума бегущей волны на основной мембране зависит от частоты звука. Таким образом, в процесс возбуждения при действии звуков разной частоты вовлекаются разные рецепторные клетки спирального органа. В улитке сочетаются два типа кодирования, или механизма различения, высоты тонов: пространственный и временной. Пространственное кодирование основано на определенном расположении возбужденных рецепторов на основной мембране. Однако при действии низких и средних тонов, кроме пространственного, осуществляется и временное кодирование: информация передается по определенным волокнам слухового нерва в виде импульсов, частота следования которых повторяет частоту звуковых колебаний (см. рис. 14.14). О настройке отдельных нейронов на всех уровнях слуховой системы на определенную частоту звука свидетельствует наличие у каждого из них специфической частотно-пороговой характеристики — зависимости пороговой интенсивности звука, необходимой для возбуждения нейрона, от частоты звуковых колебаний. Для каждого нейрона существует оптимальная, или характеристическая, частота звука, на которую порог реакции нейрона минимален, а в обе стороны по диапазону частот от этого оптимума порог резко возрастает. При надпороговых звуках характеристическая частота дает и наибольшую частоту разрядов нейрона. Таким образом, каждый нейрон настроен на выделение из всей совокупности звуков лишь определенного, достаточно узкого участка частотного диапазона. Частотно-пороговые кривые разных клеток не совпадают, а в совокупности перекрывают весь частотный диапазон слышимых звуков, обеспечивая полноценное их восприятие.
Анализ интенсивности звука. Сила звука кодируется частотой импульсации и числом возбужденных нейронов. Увеличение числа возбужденных нейронов при действии все более громких звуков обусловлено тем, что нейроны слуховой системы отличаются друг от друга по порогам реакций. При слабом стимуле в реакцию вовлекается лишь небольшое число наиболее чувствительных нейронов, а при усилении звука в реакцию вовлекается все большее число дополнительных нейронов с более высокими порогами реакций. Кроме того, пороги возбуждения внутренних и наружных рецепторных клеток неодинаковы: возбуждение внутренних волос-ковых клеток возникает при большей силе звука, поэтому в зависимости от его интенсивности меняется соотношение числа возбужденных внутренних и наружных волосковых клеток.
Слуховые ощущения. Тональность (частота) звука. Человек воспринимает звуковые колебания с частотой 16—20 000 Гц. Этот диапазон соответствует 10—11 октавам. Верхняя граница частоты воспринимаемых звуков зависит от возраста человека: с годами она постепенно понижается и старики часто не слышат высоких тонов. Различение частоты звука характеризуется тем минимальным различием по частоте двух близких звуков, которое еще улавливается человеком. При низких и средних частотах человек способен заметить различия в 1—2 Гц. Встречаются люди с абсолютным слухом: они способны точно узнавать и обозначать любой звук даже при отсутствии звука сравнения.
Слуховая чувствительность. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют абсолютным порогом слуховой чувствительности. Пороги слышимости зависят от частоты звука. В области частот 1000— 4000 Гц слух человека максимально чувствителен. В этих пределах слышен звук, имеющий ничтожную энергию. При звуках ниже 1000 и выше 4000 Гц чувствительность резко уменьшается: например, при 20 и при 20 000 Гц пороговая энергия звука в миллион раз выше (нижняя кривая AEFGD на рис. 14.16).
Усиление звука может вызвать неприятное ощущение давления и даже боль в ухе. Звуки такой силы характеризуют верхний предел слышимости (кривая ABCD на рис. 14.16) и ограничивают область нормального слухового восприятия.
Поможем написать любую работу на аналогичную тему
Реферат
Проприоцептивная сенсорная система, её роль в организации двигательного акта.
От 250 руб
Контрольная работа
Проприоцептивная сенсорная система, её роль в организации двигательного акта.
От 250 руб
Курсовая работа
Проприоцептивная сенсорная система, её роль в организации двигательного акта.
От 700 руб