В психологии внимание определяется как процесс и состояние настройки субъекта на восприятие приоритетной информации и выполнение поставленных задач.
Направленность и сосредоточенность психической деятельности при внимании обеспечивает более эффективное восприятие информации. В общем плане выделяют два основных вида внимания: непроизвольное и произвольное (избирательное, селективное). Оба вида внимания имеют разные функции, по-разному формируются в онтогенезе, и в их основе лежат различные физиологические механизмы.
1. Ориентировочная реакция
Принято считать, что физиологическую основу, на которой развивается и функционирует непроизвольное внимание, составляет ориентировочная реакция.
Ориентировочная реакция (ОР) впервые была описана И.П. Павловым как двигательная реакция животного на новый, внезапно появляющийся раздражитель. Она включала поворот головы и глаз в сторону раздражителя и обязательно сопровождалась торможением текущей условно-рефлекторной деятельности. Другая особенность ОР заключалась в угашении всех ее поведенческих проявлений при повторении стимула. Угасшая ОР, легко восстанавливалась при малейшем изменении обстановки
Физиологические показатели ОР. Использование полиграфической регистрации показало, что ОР вызывает не только поведенческие проявления, но и целый спектр вегетативных изменений. Отражением этих генерализованных изменений являются различные компоненты ОР: двигательный (мышечный), сердечный, дыхательный, кожно-гальванический, сосудистый, зрачковый, сенсорный и электроэнцефалографический. Как правило, при предъявлении нового стимула повышается мышечный тонус, изменяется частота дыхания, пульса, возрастает электрическая активность кожи, расширяются зрачки, снижаются сенсорные пороги. В электроэнцефалограмме в начале ориентировочной реакции возникает генерализованная активация, которая проявляется в блокаде (подавлении) альфа-ритма и смене его высокочастотной активностью. Одновременно с этим возникает возможность объединения и синхронной работы нервных клеток не по принципу их пространственной близости, а по функциональному принципу. Благодаря всем этим изменениям возникает особое состояние мобилизационной готовности организма.
Чаще других в экспериментах, направленных на изучение ОР, используют показатели кожно-гальванической реакции (КГР). Она обладает особой чувствительностью к новизне стимула, модальна неспецифична, т.е. не зависит от того, какой именно стимул вызывает ОР. Кроме того, КГР быстро угасает, даже если ОР вызвана болевым раздражителем. Однако КГР тесно связана с эмоциональной сферой, поэтому использование КГР при изучении ОР требует четкого разделения собственно ориентировочного и эмоционального компонентов реагирования на новый стимул.
Нервная модель стимула. Механизм возникновения и угашения ОР получил толкование в концепции нервной модели стимула, предложенной Е.Н. Соколовым. Согласно этой концепции, в результате повторения стимула в нервной системе формируется "модель", определенная конфигурация следа, в которой фиксируются все параметры стимула. Ориентировочная реакция возникает в тех случаях, когда обнаруживается рассогласование между действующим стимулом и сформированным следом, т.е. "нервной моделью". Если действующий стимул и нервный след, оставленный предшествующим раздражителем, идентичны, то ОР не возникает. Если же они не совпадают, то ориентировочная реакция возникает и оказывается до известной степени тем сильнее, чем больше различаются предшествующий и новый раздражители. Поскольку ОР возникает в результате рассогласования афферентного раздражения с "нервной моделью" ожидаемого стимула, очевидно, что ОР будет длиться до тех пор, пока существует эта разница.
В соответствии с этой концепцией ОР должна фиксироваться при любом сколько-нибудь ощутимом расхождении между двумя последовательно предъявляемыми стимулами. Имеются, однако, многочисленные факты, которые свидетельствуют о том, что ОР далеко не всегда обязательно возникает при изменении параметров стимула.
Значимость стимула. Ориентировочный рефлекс связан с адаптацией организма к меняющимся условиям среды, поэтому для него справедлив "закон силы". Иначе говоря, чем больше изменяется стимул (например, его интенсивность или степень новизны), тем значительнее ответная реакция. Однако не меньшую, а нередко и большую реакцию могут вызвать ничтожные изменения ситуации, если они прямо адресованы к основным потребностям человека.
Кажется, что более значимый и, следовательно, в чем-то уже знакомый человеку стимул должен при прочих равных условиях вызывать меньшую ОР, чем абсолютно новый. Факты, однако, говорят о другом. Значимость стимула нередко имеет решающее значение для возникновения ОР. Высоко значимый стимул может вызвать мощную ориентировочную реакцию, имея небольшую физическую интенсивность.
- По некоторым представлениям, факторы, провоцирующие ОР, можно упорядочить, выделив 4 уровня, или регистра:
o стимульный регистр;
o регистр новизны;
o регистр интенсивности;
o регистр значимости.
Первый уровень оценки проходят практически все стимулы, второй и третий регистры работают параллельно. Пройдя любой из этих двух регистров, стимул поступает в последний и там оценивается его значимость. Только после этого завершающего акта оценивания развивается весь комплекс ориентировочной реакции.
Таким образом, ОР возникает не на любой новый стимул, а только на такой, который предварительно оценивается как биологически значимый. Иначе мы переживали бы ОР ежесекундно, так как новые раздражители действуют на нас постоянно. Оценивая ОР, следовательно, надо учитывать не формальное количество информации, содержащейся в стимуле, а количество семантической, значимой информации.
Существенно и другое: восприятие значимого стимула нередко сопровождается формированием ответной адекватной реакции. Присутствие моторных компонентов свидетельствует о том, что ОР предоставляет собой единство воспринимающих и исполнительных механизмов. Таким образом, ОР, традиционно рассматриваемая как реакция на новый раздражитель, представляет частный случай ориентировочной деятельности, которая понимается как организация новых видов деятельности, формирование активности в изменившихся условиях среды
2. Нейрофизиологические механизмы внимания
Одним из наиболее выдающихся достижений нейрофизиологии в ХХ в. явилось открытие и систематическое изучение функций неспецифической системы мозга, которое началось с появления в 1949 г. книги Г. Моруцци и Г. Мэгуна "Ретикулярная формация мозгового ствола и реакция активации в ЭЭГ".
Ретикулярная формация наряду с лимбической системой образуют блок модулирующих систем мозга, основной функцией которых является регуляция функциональных состояний организма. Первоначально к неспецифической системе мозга относили в основном лишь сетевидные образования ствола мозга и их главной задачей считали диффузную генерализованную активацию коры больших полушарий. По современным представлениям, восходящая неспецифическая активирующая система простирается от продолговатого мозга до зрительного бугра (таламуса).
Функции таламуса. Таламус, входящий в состав промежуточного мозга, имеет ядерную структуру. Он состоит из специфических и неспецифических ядер. Специфические ядра обрабатывают всю поступающую в организм сенсорную информацию, поэтому таламус образно называют коллектором сенсорной информации. Специфические ядра таламуса связаны, главным образом, с первичными проекционными зонами анализаторов. Неспецифические ядра направляют свои восходящие пути в ассоциативные зоны коры больших полушарий. В 1955 г. Г. Джаспером было сформулировано представление о диффузно-проекционной таламической системе. Опираясь на целый ряд фактов, он утверждал, что диффузная проекционная таламическая система (неспецифический таламус) в определенных пределах может управлять состоянием коры, оказывая на нее как возбуждающее, так и тормозное влияние.
В экспериментах на животных было показано, что при раздражении неспецифического таламуса в коре головного мозга возникает реакция активации. Эту реакцию легко наблюдать при регистрации энцефалограммы, однако активация коры при раздражении неспецифического таламуса имеет рад отличий от активации, возникающей при раздражении ретикулярной формации ствола мозга.
Таблица 6.1.
Реакции активации структур мозга
|
Характеристики реакции |
ЭЭГ активация ствола мозга |
ЭЭГ активация таламуса |
|
Область распространения |
Генерализованная |
Локальная |
|
Тип реакции |
Тоническая |
Фазическая |
|
Временная динамика |
Медленно угасающая |
Быстро угасающие |
- По современным представлениям, переключение активирующих влияний с уровня ретикулярной формации ствола мозга на уровень таламической системы означает переход от генерализованной активации коры к локальной:
1. Первая отвечает за глобальные сдвиги общего уровня бодрствования;
2. Вторая отвечает за избирательное сосредоточение внимания.
Функции фронтальных зон. Ретикулярная формация ствола мозга и неспецифический таламус тесно связаны с корой больших полушарий. Особое место в системе этих связей занимают фронтальные зоны коры. Предполагается, что возбуждение ретикулярной формации ствола мозга и неспецифического таламуса по прямым восходящим путям распространяется на передние отделы коры. При достижении определенного уровня возбуждения фронтальных зон по нисходящим путям, идущим в ретикулярную формацию и таламус, осуществляется тормозное влияние. Фактически здесь имеет место контур саморегуляции: ретикулярная формация изначально активизирует фронтальную кору, а та в свою очередь тормозит (снижает) активность ретикулярной формации. Поскольку все эти влияния носят градуальный характер, т.е. изменяются постепенно, то с помощью двухсторонних связей фронтальные зоны коры могут обеспечивать именно тот уровень возбуждения, который требуется в каждом конкретном случае.
Таким образом, фронтальная кора — важнейший регулятор состояния бодрствования в целом и внимания как избирательного процесса. Она модулирует в нужном направлении активность стволовой и таламической систем. Благодаря этому можно говорить о таком явлении, как управляемая корковая активация.
Система внимания в мозге человека. Изложенная выше схема не исчерпывает всех представлений о мозговом обеспечении внимания. Она характеризует общие принципы нейрофизиологической организации внимания и адресуется, главным образом, к так называемому модально-неспецифическому вниманию. Более детальное изучение позволяет специализировать внимание, выделив его модально-специфические виды. Как относительно самостоятельные можно описать следующие виды внимания: сенсорное (зрительное, слуховое, тактильное), двигательное, эмоциональное и интеллектуальное. Клиника очаговых поражений показывает, что эти виды внимания могут страдать независимо друг от друга и в их обеспечении принимают участие разные отделы мозга. В поддержании модально-специфических видов внимания принимают активное участие зоны коры, непосредственно связанные с обеспечением соответствующих психических функций).
Известный исследователь внимания М. Познер утверждает, что в мозге человека существует самостоятельная система внимания, которая анатомически изолирована от систем обработки поступающей информации. Внимание поддерживается за счет работы разных анатомических зон, образующих сетевую структуру, и эти зоны выполняют разные функции, которые можно описать в когнитивных терминах. Причем выделяется ряд функциональных подсистем внимания. Они обеспечивают три главные функции: ориентацию на сенсорные события, обнаружение сигнала для фокальной (сознательной обработки) и поддержание бдительности, или бодрствующего состояния. В обеспечении первой функции существенную роль играет задняя теменная область и некоторые ядра таламуса, второй — латеральные и медиальные отделы фронтальной коры. Поддержание бдительности обеспечивается за счет деятельности правого полушария.
Действительно, немало экспериментальных данных свидетельствует о разном вкладе полушарий в обеспечение не только восприятия, но и избирательного внимания. По этим данным, правое полушарие в основном обеспечивает общую мобилизационную готовность человека, поддерживает необходимый уровень бодрствования и сравнительно мало связано с особенностями конкретной деятельности. Левое в большей степени отвечает за специализированную организацию внимания в соответствии с особенностями задачи.
3. Методы изучения и диагностики внимания
Экспериментальное изучение физиологических коррелятов и механизмов внимания осуществляется на разных уровнях, начиная от нервной клетки и кончая биоэлектрической активностью мозга в целом. Каждый из этих уровней исследования формирует свои представления о физиологических основах внимания.
Нейроны новизны. Наиболее интересные факты, иллюстрирующие функции нейронов в механизмах внимания, связаны с обеспечением ориентировочной реакции. Еще в 60-е гг. Г. Джаспер во время нейрохирургических операций выделил в таламусе человека особые нейроны — "детекторы" новизны, или внимания, которые реагировали на первые предъявления стимулов.
Позднее в нейронных сетях были выделены нервные клетки, получившие название нейронов новизны и тождества. Нейроны новизны позволяют выделять новые сигналы. Они отличаются от других характерной особенностью: их фоновая импульсация возрастает при действии новых стимулов разной модальности. С помощью множественных связей эти нейроны соединены с детекторами отдельных зон коры головного мозга, которые образуют на нейронах новизны пластичные возбуждающие синапсы. Таким образом, при действии новых стимулов импульсная активность нейронов новизны возрастает. По мере повторения стимула и в зависимости от силы возбуждения ответ нейрона новизны избирательно подавляется, так, что дополнительная активация в нем исчезает и сохраняется лишь фоновая активность.
Нейрон тождества также обладает фоновой активностью. К этим нейронам через пластичные синапсы поступают импульсы от детекторов разных модальностей. Но в отличие от нейронов новизны, в нейронах тождества связь с детекторами осуществляется через тормозные синапсы. При действии нового раздражителя фоновая активность в нейронах тождества подавляется, а при действии привычных раздражителей, напротив, активизируется.
Итак, новый стимул возбуждает нейроны новизны и тормозит нейроны тождества, таким образом новый раздражитель стимулирует активирующую систему мозга и подавляет синхронизирующую (тормозную) систему. Привычный стимул действует прямо противоположным образом — усиливая работу тормозной системы, не влияет на активирующую.
Особенности импульсной активности нейронов человека при выполнении психологических проб, требующих мобилизации произвольного внимания, описаны в работах Н.П. Бехтеревой и ее сотрудников. При этом в передних отделах таламуса и ряде других структур ближайшей подкорки были зафиксированы стремительные возникающие вспышки импульсной активности, по частоте в 2-3 раза превышающие уровень фона. Характерно, что описанные изменения в импульсной активности нейронов сохранялись на протяжении выполнения всего теста, и только по его завершении уровень активности этих нейронов возвращался к исходному.
В целом в этих исследованиях установлено, что различные формы познавательной деятельности человека, сопровождающиеся напряжением произвольного внимания, характеризуются определенным типом нейрональной активности, четко сопоставимым с динамикой произвольного внимания.
Электроэнцефалографические корреляты внимания. Хорошо известно, что при предъявлении стимула в энцефалограмме наблюдается подавление (блокада) альфа-ритма, и на смену ему приходит реакция активации. Однако этим не исчерпываются изменения электрической активности мозга в ситуации внимания.
Исследование суммарной электрической активности при мобилизации интеллектуального внимания выявило закономерные изменения в характере совместной деятельности разных зон коры. При оценке степени дистантной синхронизации биопотенциалов было установлено, что в передних зонах левого полушария существенно по сравнению с фоном увеличивается уровень пространственной синхронизации. Сходные результаты дает использование и другого показателя, извлекаемого из энцефалограммы, — когерентности. В ситуации ожидания стимула независимо от его модальности наблюдается рост когерентности в полосе альфа-ритма, причем преимущественно в передних (премоторных) зонах коры. Высокие показатели дистантной синхронизации и когерентности говорят о том, насколько тесно взаимодействуют зоны коры, в первую очередь передних отделов левого полушария, в обеспечении произвольного внимания.
Изучение внимания с помощью ВП. Первые исследования внимания методом ВП использовали простые поведенческие модели, например, счет стимулов. При этом было установлено, что привлечение внимания испытуемых к стимулу сопровождается увеличением амплитуды компонентов ВП и сокращением их латентности. Напротив, отвлечение внимания от стимула сопровождается снижением амплитуды ВП и увеличением латентности. Однако оставалось неясным, чем обусловлены эти изменения параметров ВП: изменением общего уровня активации, поддержанием бдительности или механизмами избирательного внимания. Для разведения указанных процессов необходимо было построить эксперимент таким образом, чтобы его организация позволяла вычленить эффект мобилизации селективного внимания "в чистом" виде.
В качестве такой модели можно привести эксперименты С. Хильярда, которые получили в 70-е гг. широкую известность. При предъявлении звуковых стимулов через наушники в левое и правое ухо испытуемому предлагается мысленно реагировать (считать) редко встречающиеся ("целевые") стимулы, поступающие по одному из каналов (только в правое или левое ухо). В результате получают вызванные потенциалы в ответ на 4 варианта стимулов: часто встречающиеся в релевантном (контролируемом) и иррелевантном (игнорируемом) каналах и редко встречающиеся (целевые) в том и другом каналах. В этом случае появляется возможность сравнивать эффекты канала и стимула, которые являются объектом внимания. В экспериментах такого типа, как правило, применяются очень короткие интервалы между стимулами (немногим более или менее одной секунды), в результате усиливается напряженность и устойчивость избирательного внимания испытуемого к быстро чередующимся стимулам разной информационной значимости.
|
|
Слуховые вызванные потенциалы, отражающие привлечение селективного внимания к одному из каналов в ситуации различения звуковых сигналов (700 или 300 Гц) (по H. Hansen & S. Hillyard, 1982). |
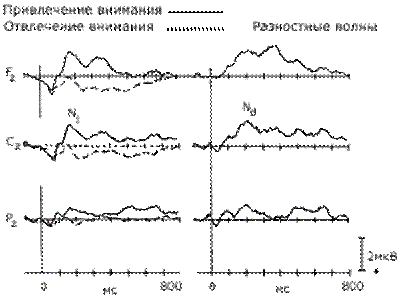
Было установлено, что привлечение внимания к одному из каналов ведет к увеличению амплитуды первой отрицательной волны с латентным периодом около 150 мс, обозначаемой как компонент N1. Целевые стимулы сопровождались появлением в составе ВП позднего положительного колебания Р3 с латентным периодом около 300 мс. Высказывалось предположение, что негативная волна N1 отражает "установку" на стимул, определяющую направленность произвольного внимания, а компонент Р3 — "установку на ответ", связанную с выбором варианта ответа. В дальнейшем компонент Р3 (чаще определяемый как Р300) явился предметом множества исследований.
В более поздних исследованиях с помощью специального приема вычитания потенциалов, регистрируемых в ответ на сигнальные и стандартные стимулы, обнаружилось, что первая отрицательная волна N1 представляет собой неоднородный корковый феномен сложной структуры, в котором можно выделить особое отрицательное колебание, так называемую — "негативность, отражающую обработку информации". Это колебание с латентным периодом около 150 мс и длительностью не менее 500 мс регистрируется при несовпадении редко предъявляемого целевого  стимула со "следом внимания", образуемым в ассоциативной слуховой зоне и лобной области при частом повторении и воспроизведении стандартного стимула. При этом, чем меньше разница между этими стимулами, тем больше латентный период и тем длительнее отрицательное колебание, развивающееся в ответ на целевой, нестандартный стимул.
стимула со "следом внимания", образуемым в ассоциативной слуховой зоне и лобной области при частом повторении и воспроизведении стандартного стимула. При этом, чем меньше разница между этими стимулами, тем больше латентный период и тем длительнее отрицательное колебание, развивающееся в ответ на целевой, нестандартный стимул.
Кроме этого, описано еще одно отрицательное колебание, в ряде случаев сопровождающее ситуацию сравнения стимулов. Этот компонент, обозначаемый как "негативность рассогласования", возникает в слуховой коре с латентным периодом 70-100 мс и отражает автоматический процесс сравнения физических признаков звукового стимула со следом стандартного стимула, хранящемся в течение 5-10 сек в сенсорной памяти. При отклонении физических свойств стимула от следа многократно предъявляемого стандартного стимула развивается "негативность рассогласования".
Предполагается, что в образовании волны N1 могут участвовать оба компонента ("негативность, связанная с обработкой информации" и "негативность рассогласования"). Причем первый из этих компонентов связан с предсознательной, непроизвольной оценкой признаков необычного звукового стимула, осуществляемой путем сравнения их с нервной моделью часто повторяющегося стимула, а второй компонент отражает процессы обработки сенсорной информации на сознательном уровне, а именно: произвольного внимания, фокусирования субъектом сознания на определенных критических признаках стимула и сравнения его со "следом внимания", хранящемся в рабочей памяти.
Таким образом, с помощью метода ВП было показано, что на целевые звуковые стимулы (в ситуации выбора стимула и канала) возникает два типа компонентов, один из которых отражает процессы сенсорной памяти, другой — селективного внимания.
Временные характеристики внимания. С помощью метода ВП можно оценить динамику развития процессов внимания в реальном времени. Вопрос заключается в следующем, на каком этапе обработки информации включаются процессы внимания? Поскольку начало первой негативной волны, возникающей в ответ на сигнальные стимулы, в основном приурочено к 50 мс от момента предъявления стимула, пятидесятимиллисекундная граница довольно долго рассматривалась как временной рубеж, после которого развертываются процессы селективного внимания.
Более детальные исследования, однако, показали, что в слуховой и, по-видимому, соматосенсорной системах произвольная регуляция процессов обработки поступающей информации включается не позже, чем через 20-30 сек. после предъявления стимула. Эффекты внимания в зрительной системе обнаруживают себя позднее, начиная с 60 мс. Не исключено, что и эти временные границы по мере совершенствования методов изучения будут изменены. Суть, однако, в том, что хронометрия переработки информации и включения внимания как одного из главных регуляторов этого процесса с такой точностью может быть изучена только в психофизиологических экспериментах.
Поможем написать любую работу на аналогичную тему