Предки покрытосеменных, как указывалось ранее, остаются до настоящего времени неизвестными. Но, несомненно, вся эта группа возникла от голосеменных и обособилась от них, вероятно, еще на ранних этапах всей истории семенных растений. Из современных покрытосеменных, соответственно стробилярной теории цветка, по-видимому, наиболее близок к древнейшим покрытосеменным подкласс Magnoludae .Однако Magnolndae имеют уже настолько специализированный аппарат для привлечения насекомых-опылителей, что естественно предположить существование в прошлом еще более древней группы покрытосеменных. Такие гипотетические предковые покрытосеменные могли быть неустойчиво энтомофильными растениями, у которых перенос пыльцы совершался и ветром ,насекомыми. По всей вероятности, у них достаточно широко было распространено и самоопыление.
Рисунок 1
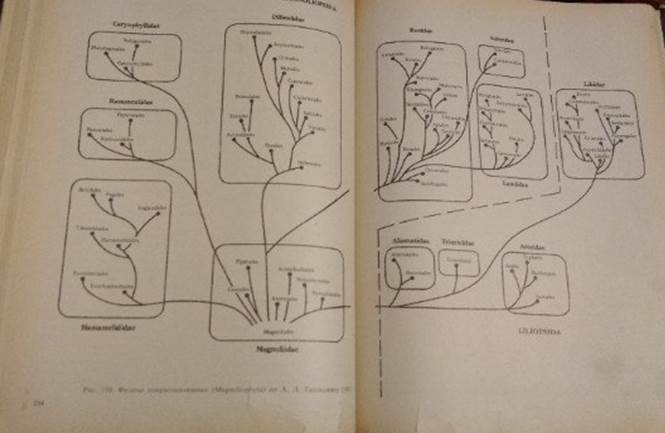
Схема эволюции цветковых растений
Гипотетические покрытосеменные предки современных цветковых дали начало, вероятно, нескольким линиям эволюции . Главной из них, т. е. имевшей впоследствии наибольший успех в освоении самых разнообразных условий жизни, была линия, в которой рано выработались Magnoliidae. Другой была линия, представленная подклассом Hamamelidae (см.рис.)
В конце прошлого — начале текущего столетия было широко распространено представление об однопокровных, которые в основном соответствуют Hamamelidae, как о группе весьма древней, примитивной, близкой к предкам покрытосеменных.Учение о происхождении покрытосеменных от голосеменных через однопокровные дополнялось специально разработанной австрийским ботаником Р.Веттштейном теорией происхождения цветка покрытосеменных из "соцветия" голосеменных (типа эфедры). Эта теория получила название теории псевданция (ложного цветка).
Научная критика концепции происхождения всего многообразия покрытосеменных от однопокровных вскрыла прежде всего, что морфологически "простое" строение цветка может быть следствием упрощения, возникшего в связи с особым направлением отбора. Это подтверждается наличием остаточного околоцветника (дуб, лещина), появлением в цветках рудиментов недостающего пола и развитием в качестве исключения обоеполых цветков вместо однополых.
Если простота цветков однопокровных вторичная, то приходится заключить, что эта группа прошла длительный путь эволюции, возникнув от предков с обоеполыми цветками. Но обоеполость цветков тесно связана с энтомофилией. Поэтому и распространенная у однопокровных анемофилия должна была прийти на смену энтомофилии, что возможно в процессе приспособления к жизни большими одновидовыми скоплениями.По-видимому, именно благодаря этому от каких-то энтомофильных лилиецветных возникли анемофильные осоки и злаки; анемофильные полыни, несомненно, являются производными энтомофильных сложноцветных.
Подвергалась глубокой критике и псевданциевая теория происхождения цветка, оказавшаяся неубедительной.В итоге, на смену представлению о первичности и примитивности однопокровных сформировалось прямо противоположное воззрение — однопокровных стали рассматривать как группу, прошедшую длительный путь эволюции и возникшую от высокоспециализированных покрытосеменных.При этом допускалось, что различные порядки однопокровных — производные разных эволюционных линий покрытосеменных. Это означало отрицание группы как единого естественного таксона.Схематический обзор родственных связей покрытосеменных растений позволяет сделать несколько обобщающих выводов. Ход эволюционного развития осуществлялся в виде нескольких параллельных потоков, которые соответствуют подклассам и классам.
Таким образом, система связей между покрытосеменными не может быть уподоблена родословному дереву; она, скорее, соответствует "кустарнику" на том этапе его развития, когда он уже потерял главный корень. Но в таком случае приходится признавать, что однокоренное, монофилетическое происхождение покрытосеменных продолжает оставаться гипотезой, хотя и очень вероятной. Гипотетичность монофилетического происхождения покрытосеменных усугубляется и тем, что пока нет возможности безоговорочно указать группу древних растений, вероятнее всего голосеменных, которая дала начало первым цветковым. При таких обстоятельствах не приходится удивляться, что по временам возникали и продолжают появляться гипотезы о полифилетическом возникновении цветковых. Однако полифилетические воззрения находятся не в лучшем положении, так как выдвигают задачу отыскания не одного, а нескольких предков покрытосеменных, что не смягчает, а обостряет трудность проблемы.
В схеме родственных отношений покрытосеменных растений достаточно четко отражен параллелизм в развитии подклассов и даже классов. Одно из его проявлений состоит в том, что почти в каждом подклассе имеются энтомофильные и анемофильные ветви. Распространенность анемофильных приспособлений в разных эволюционных потоках свидетельствует о том, что существуют какие-то общие условия, при которых анемофилия представляет существенное преимущество в борьбе за существование. Одно из таких условий — способность произрастания массовыми одновидовыми скоплениями, в которых опыление ветром, при производстве большого количества пыльцы, перестает быть ненадежным. Но произрастание в форме массовых скоплений возможно при условии,что внутривидовая конкуренция становится инструментом саморегуляции численности, поддерживающим ее на уровне, необходимом для подавления конкурентов. С этой точки зрения устойчивое ветроопыление, если оно обеспечивается надежными микробиологическими особенностями цветков и соцветий, представляет высокое достижение в ходе исторического развития цветковых.Но все же более распространенным у покрытосеменных оказалось опыление животными: насекомыми, птицами, рукокрылыми.
Почти в каждом подклассе можно найти растения (главным образом зоофильные) с правильными и неправильными (зигоморфными) цветками, с раздельнолепестным венчиком, с верхней и нижней завязью. На основании морфогенеза, спайнолепестности, зигоморфности и нижней завязи можно с уверенностью сказать, что эти признаки являются более поздними, вторичными по сравнению с первичными: раздельнолепестностью и актиноморфностью цветка, верхней завязью. Эти данные, принимая во внимание приспособительное значение вторичных признаков, помогают филогенетику размещать порядки и семейства в пределах эволюционных рядов соответственно формированию во времени. Однако факт, что указанные вторичные морфобиологические особенности имеют место в энтомофильной эволюции во всех подклассах, приходится рассматривать как закономерность. Нет необходимости видеть причину этого во "внутренних свойствах" растений, "предопределяющих", как иногда думают, эволюционные изменения. Возможности изменений органов у растений в целом очень ограничены.
Сдвиги в онтогенезе цветка в сторону более раннего наступления базального роста у оснований примордиев лепестков, что приводит к возникновению спайнолепестности, — достаточная основа для последующего отбора, поскольку трубка венчика ограничивает круг потребителей продуктов, вырабатываемых цветком, и одновременно защищает эти продукты от посетителей, которые не могут произвести опыление. Аналогичное можно сказать и о возникновении других вторичных особенностей цветка.
Основные контуры эволюционных рядов покрытосеменных растений были построены главным образом на основе учета разнообразия цветков и плодов. В настоящее время эти ряды или те, которые получены в результате критического пересмотра ранее построенных, все чаще находят подтверждение в особенностях вегетативного тела. В этом отношении очень важные результаты были получены при анатомическом изучении покрытосеменных растений, в частности их водопроводящей системы. .
Настоящие сосуды, как известно, свойственны только покрытосеменным. Если подобные образования и встречаются в более древних группах (голосеменные, папоротникообразные), то это только аналоги сосудов. Но членики сосудов покрытосеменных различны. Среди них можно отличать и более типичные, и приближающиеся к трахеидам. Кроме того, известно, что среди покрытосеменных есть и бессосудистые растения. Такие группы естественно рассматривать как более древние. В некоторых эволюционных рядах, отвечающих прогрессивному изменению цветка, обнаруживается последовательное нарастание признаков совершенствования водопроводящей аппаратуры растения. Подтверждаются эти ряды и другими анатомическими признаками.
В течение последних десятилетий большого прогресса достигла особая ветвь морфологии — морфология пыльцы. Ее данные, привлекаемые к построению филогенетических рядов, помогают ориентироваться в высоте организации, а также в наличии сходств, которые можно истолковывать как показатели родства.Хотя представления о ходе эволюционного развития, показанные прилагаемым схематическим рисунком, нельзя считать окончательным, все же в основном они правильно отражают, во-первых, взаимосвязь по крайней мере более крупных подразделений покрытосеменных, а во-вторых, размещение в каждом из стволов более поздних (по возникновению в пределах данной филогенетической ветви) и более ранних таксонов.
Эта схема позволяет выделить в многообразии покрытосеменных три уровня. Первый, наиболее низкий уровень включает примитивных покрытосеменных, их неизвестных предков и обширный подкласс Magnoliidae. Этому уровню противоположен другой, наивысший, характеризуемый спайнолепестностью и часто зигоморфностью цветка, нижней завязью, тенденцией к мельчанию цветков и образованию компактных соцветий. Этот уровень достаточно ясен у энтомо- фильных двудольных. У высших энтомофильных однодольных соответственно наблюдается зигоморфность цветков, сопровождаемая порой глубокими перестройками андроцея и гинецея, а также появление сложных аппаратов, обеспечивающих анемофилию. Между этими двумя уровнями находит себе место промежуточный, в котором отразилось много частных проявлений эволюционного процесса.
Таблица 1
Охватывающая эволюцию растений по геологическим периодам
|
Эра |
Геологический период |
Млн.лет |
Преобладающие/распространенные растения |
|
Кайнозой |
Третичный и четвертичный |
До 65 |
Преобладание/распространение покрытосеменных |
|
Меловой |
До 144 |
Преобладание покрытосеменных |
|
|
Мезозой |
Юрский |
До 213 |
Доминируют голосеменные и птеридофиты (хвойные деревья, папоротники, хвощи, плауны). |
|
Триасовый |
До 248 |
Распространение голосеменных. Пышные леса. |
|
|
Пермский |
До 286 |
Преобладают птеридофиты, или примитивные сосудистые растения (папоротники, хвощи, плауны). Распространены хвойные деревья и гинкго. |
|
|
Каменноугольный |
До 360 |
Птеридофиты доминируют в углеобразующих болотах. |
|
|
Палеозой |
Девонский |
До 408 |
Птеридофиты доминируют в углеобразующих болотах. |
|
Силурийский |
До 440 |
Появляются первые наземные/болотные растения. |
|
|
Ордовикский |
До 550 |
Водоросли. |
|
|
Кембрийский |
До 590 |
Водоросли. |
|
|
Докембрийский |
Более 590 |
Сине-зелёные водоросли. |
Поможем написать любую работу на аналогичную тему