В чем состоят проблемы моделирования ГС адаптивного признака? Реальная сеть обычно обладает очень сложной, иерархической структурно-функциональной организацией, нередко содержащей сотни компонентов. Можно выделить несколько уровней организации ГС, которые учитываются при моделировании этих объектов.
Самым нижним является биохимический уровень. Здесь рассматривается огромная сеть биохимических процессов, происходящих в клетке или организме. Следующим уровнем является генетический. Здесь уже учитывается расположение и ориентация генов, регуляторных элементов, полиаллельность и т.д. Затем следует выделить клеточный уровень, где учитываются процессы активного и пассивного переноса, выделить зоны (компартменты) локализации и действия ГС и т.д. Также выделяют более высокие уровни организации – тканевый, органный, организменный, популяционный и т.д. В большинстве случаев, объекты моделирования обладают сложным нелинейным поведением в силу наличия в них отрицательных и положительных ОС.
Даже если ограничиться рассмотрением ГС на уровне отдельной клетки, можно выделить несколько зон, например, межклеточное пространство, клеточная мембрана, цитоплазма, ядро. Кроме общих, в каждой зоне идут свои процессы, продукты которых воздействуют на ГС других зон. Далее необходимо описать ГС, отдельные части которых распределены по более крупным компартментам организма, таким, как группы клеток, органы и ткани. Для полного описания генных сетей желательно рассмотрение протекающих в них взаимосвязанных процессов на уровне целого организма.
При этом возникают трудности, связанные как с моделированием систем очень большой размерности, так и проведением экспериментов с реальными ГС. Поэтому пока исследования концентрируются на относительно небольших ГС, допускающих прямое сравнение с экспериментальными данными.
Базы данных
Для построения модели любой ГС используют информацию об ее элементах и связях между ними. Это означает, что необходимо знать какие гены участвуют в формировании сети, их регуляторные элементы (промоторы и др.), синтезируемые ими белки, пути передачи сигналов в клетке. Привлечение этой информации идет путем обобщения литературных данных и получения новых данных в ходе натурных экспериментов с конкретными ГС. Далее, составляют уравнения, характеризующие динамику ГС, и подбирают значения параметров – констант, входящих в эти уравнения.
Основной проблемой стало отсутствие эффективных компьютерных технологий для ведения информации об их структурно-функциональной организации, прежде всего для эукариот. С целью накопления и обобщения информации создан целый ряд баз данных . В них содержится качественная, описательная информация (о структурно-функциональной организации генной сети, механизмы различных процессов, организация регуляторных участков генов и т.д.), а также количественная информация (параметры ферментативных реакций, массы белков, и т.д.).
В качестве примера отметим базу данных GeneNet, созданную в ИЦиГ и доступную через интернет . Она является частью системы GeneExpress. Классификация компонентов ГС в БД GeneNet представлена на рис. 5.
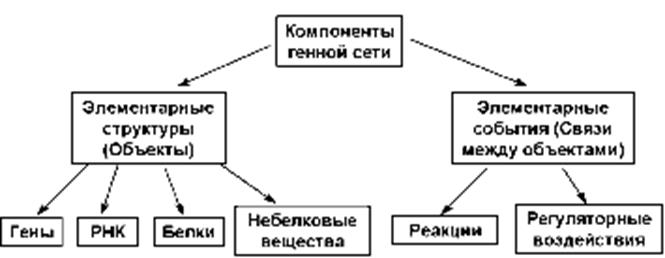
Рисунок 5 – Классификация компонентов генной сети в системе GeneNet
Для моделирования ГС в GeneNet разработан специальный язык и удобный графический интерфейс. Можно, в частности, указать элемент модели ГС, изменить его характеристики и проследить на графике ответные изменения в поведении различных компонентов ГС.
Технология GeneNet позволяет эксперту реконструировать ГС на основе экспериментальных данных, опубликованных в научных статьях. В базе данных описано несколько сотен генов, белков и небелковых веществ 43 организмов по данным из сотен публикаций. В результате удалось построить модели некоторых ГС: липидного метаболизма, кроветворения, противовирусного ответа, функционирования эндокринной системы, регуляции развития семени, азотфиксации, теплового шока, регуляции холестерина и др.
Графические схемы функционирования генных сетей
Генные сети в упрощенных вариантах моделей описывают на языке генетических элементов (ГЭ). Каждый из них содержит ген, соответствующие ему мРНК и белки. Функционирование генетического элемента в простейшем случае – процесс синтеза белка. В генетический элемент неявно включаются все процессы, обеспечивающие синтез мРНК, белка, формирование мультимера белка (мономерная форма белка есть частный случай мультимерной). Белки обеспечивают регулирующие взаимодействия между ГЭ.
Регуляторный механизм ГС – элементарный процесс, посредством которого некоторая совокупность генетических элементов действует на конкретный генетический элемент, уменьшая или увеличивая скорость синтеза его продукта. В реальной ГС кроме белков генетические элементы, возможно, содержат низкомолекулярные соединения, которые также регулируют состояния других ГЭ.
Схему функционирования ГС, включающую ГЭ и их воздействия, можно задать ориентированным графом. Он состоит из конечного множества вершин (gi) и набора упорядоченных их пар, изображаемых в виде стрелок. Любая стрелка называется дугой, или ориентированным ребром. Она отражает направление и характер воздействия (отрицательное или положительное регуляторное воздействие) одного ГЭ на другой. Пример схемы функционирования простой гипотетической ГС – на рис. 6. Отметим, что ген 1 не регулирует экспрессию гена 2 непосредственно, но может повлиять на нее через ген 3. В некоторых случаях отрицательные регуляторные связи на схемах функционирования ГС обозначают пунктирными, а положительные – сплошными стрелками.
а) б)

Рисунок 6 – а) Исходная ГС из трех генов (внизу), их белков (наверху) и регуляторных воздействий (стрелки) на другие гены; б) соответствующий ей ориентированный граф – структурная схема функционирования ГС.
На рис. 7 в этих терминах, но в других обозначениях, показан фрагмент схемы реальной ГС прорастания семени растения. Элементы гиббереллины (gibberelins) и абсцизовая кислота (ABА) соответствуют низкомолекулярным соединениям, остальные ГЭ – гены и их белки. Стрелки, помеченные кружками с плюсом – положительные (усиливающие) регуляторные воздействия, с минусом – отрицательные (ингибирующие).
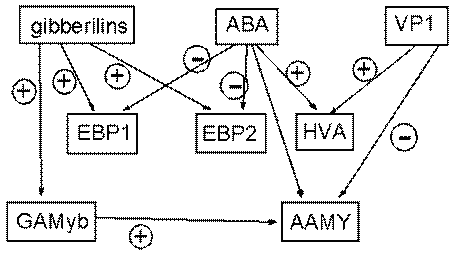
Рисунок 7 – Граф фрагмента внутриклеточной ГС прорастания семени
Графическое моделирование – простейший способ описания и первичного анализа структурно-функциональной организации ГС. В некоторых случаях непосредственно по структурному графу можно выявить важные регуляторные контуры, мотивы, а также предсказывать свойства ГС, связанные с предельными режимами ее функционирования. Например, в некоторых случаях удается оценить число точек покоя или предельных циклов.
В других случаях непосредственно по графу ГС удается выявить сочетание из двух или большего числа продуктов синтеза, которые даже при длительном функционировании будут присутствовать в ГС. Такие сочетания называются предельными циклами, т.к. их концентрации продолжают циклически меняться во времени. Однако по графу ГС невозможно оценить закономерности изменения концентраций во времени, для этого их необходимо дополнить непрерывными моделями.
Таким образом, графическое моделирование используется совместно с более сложными математическими моделями.
Дискретный способ моделирования генных сетей
Существует два основных количественных детерминистических подхода к описанию ГС – дискретный и непрерывный. В некоторых случаях эти математические модели объединяют, а также пытаются учесть случайную составляющую процессов в ГС с помощью стохастических моделей. Последние применяют, в частности, для таких генных сетей, в которых значительные события обеспечиваются очень малым количеством регуляторного агента (например, транскрипционного фактора).
В простейших вариантах дискретных моделей ГС предполагается, что каждый ген в ГЭ может находиться лишь в одном из двух состояний: 1- активен (экспрессируется) и 0 - неактивен. Для простоты предполагается, что процесс функционирования ГС состоит из элементарных этапов (шагов). На очередном шаге состояние каждого гена сохраняется или меняется на противоположное. Это зависит от активизирующих и ингибирующих воздействий на него со стороны других генов, активных перед очередным шагом. Исчерпывающей характеристикой ГС в рамках подобной модели является вектор состояний. Это набор нулей и единиц, упорядоченный по номерам ГЭ, представленных на структурном графе ГС.
Алгоритм определения состояния каждого ГЭ по шагам формализован в компьютерной программе. Для этого используют специальные функции, оперирующие только с 0 и 1 (алгебра логики – булева алгебра), или пороговые функции.
Главным преимуществом дискретных моделей ГС является их простота. Такие модели позволяют прослеживать динамику ГС (изменение вектора состояний) в течение длительного времени, прогнозировать предельные режимы функционирования регуляторных контуров генных сетей. Но, безусловно, подобные модели являются довольно грубым упрощением.
Непрерывные модели
Наиболее сложная задача – изучение динамики количественных переменных ГС. Сюда относятся концентрации продуктов: «входных» - действующих на ГС извне, «внутренних» - регуляторов процессов и «выходных» - выделяемых сетью во вне. Для количественного описания динамики этих непрерывных количественных переменных во времени необходимы сложные модели. Наиболее полное описание обеспечивают дифференциальные уравнения и их системы, хотя этот подход имеет ряд недостатков.
С помощью непрерывных моделей пытаются ответить на некоторые вопросы теории ГС. Среди них:
- выявление условий существования в ГС множества состояний устойчивых во времени, т.е. состояний, равновесных по значениям основных переменных, а также выявление условий переключения между такими состояниями.
- изучение «степени устойчивости» положений равновесия и влияния на нее различных факторов (мутаций, гетерозиготности, стрессов и пр.). Понятно, например, что наличие отрицательной ОС увеличивает, а положительной - снижает устойчивость равновесия.
- нахождение условий возникновения колебательных, циклических режимов функционирования ГС. В частности, оценка минимального уровня сложности сети и необходимости присутствия в ней положительных и отрицательных ОС.
Кроме теоретических исследований ведутся работы по математическому моделированию реальных ГС и сравнению результатов с данными экспериментов.
Для построения непрерывных моделей требуется формализовать, например, с помощью дифференциальных уравнений «элементарные события» ГС: функционирование регуляторных механизмов, связанных с управлением процессами транскрипции, трансляции, биохимические реакции, включая формирование белков, передачу внешних и внутренних сигналов, работу контуров управления и пр.
В рамках непрерывных и дискретных моделей возможен гибридный подход к моделированию динамики ГС. В нем разделяют сеть на 2 подсистемы. Первая – регуляторная, включает совокупность регуляторных генов и их макромолекулы. Ее состояния моделируются на языке дискретных моделей (например, с использованием пороговых функций). Вторая – исполняющая, содержит гены, обеспечивающие выполнение различных биохимических процессов. Она моделируется системами дифференциальных уравнений. В итоге общая модель формулируется на языке кусочно-непрерывных систем уравнений. Частные примеры подобных элементарных событий и уравнений представлены на рис. 8-10.
 |
|
![]()
![]()
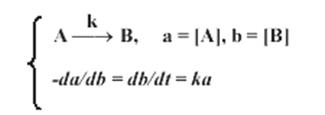
Рисунок 8 – Схема и дифференциальные уравнения – описание химико-кинетической закономерности простейшей мономолекулярной биохимической реакции фосфорилирования. Здесь a и b – концентрации исходного и результирующего продуктов реакции, k – параметр, характеризующий скорость прохождения реакции .
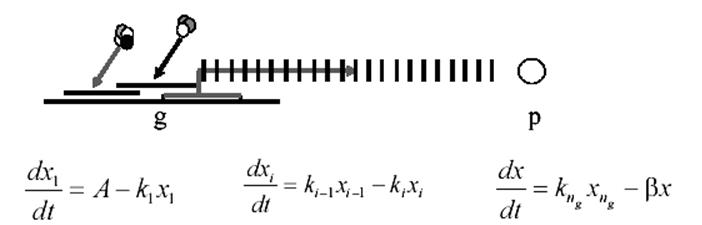
Рисунок 9 – Схема и дифференциальные уравнения, описывающие функционирование генетического элемента сети при многоэтапном формировании регуляторного белка р.
Здесь: ng – количество промежуточных стадий синтеза белка, xi, – концентрации промежуточных продуктов синтеза белка, x - концентрация белка, А – параметр - скорость инициации конститутивного синтеза, ki – параметры - скорость перехода из i-го промежуточного состояния в (i+1)-ое промежуточное состояние, β – параметр - скорость деградации белка p.
а б
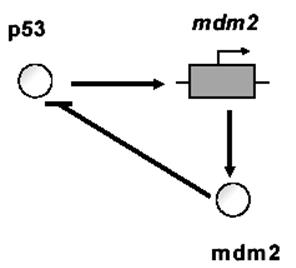
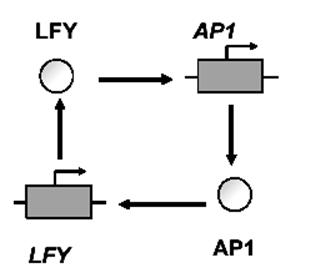
Рисунок 10 – Схема некоторых контуров ГС с обратной связью
а) Отрицательная ОС: транскрипционный фактор гена р53 активирует транскрипцию его гена-ингибитора mdm2. Уравнение для переменной x – концентрации транскрипционного фактора гена имеет вид: dx/dt=α/(1+Kc xc). α, K, с – параметры реакций. Скорость синтеза белка (dx/dt) убывает с ростом концентрации x.
б) Положительная ОС: взаимное усиление транскрипции двух генов. Уравнение для переменной x – концентрации транскрипционного фактора гена AP1 имеет вид: dx/dt=α/(1+Kc1xc)/(1+Kd2xd). Скорость синтеза белка (dx/dt) возрастает с ростом концентрации x.
В рамках непрерывных и дискретных моделей возможен гибридный подход к моделированию динамики ГС. В нем разделяют сеть на 2 подсистемы. Первая – регуляторная, включает совокупность регуляторных генов и их макромолекулы. Ее состояния моделируются на языке дискретных моделей (например, с использованием пороговых функций). Вторая – исполняющая, содержит гены, обеспечивающие выполнение различных биохимических процессов. Она моделируется системами дифференциальных уравнений. В итоге общая модель формулируется на языке кусочно-непрерывных систем уравнений.
Подобные принципы построения моделей функционирования ГС заложены в обобщенном химико-кинетическом методе моделирования (ОХКММ), разработанном в ИЦиГ. Вначале проводится иерархическая декомпозиция исходной ГС до элементарных структур и процессов. Затем процессы формализуются в терминах стандартных блоков – систем уравнений. Из них проводят сборку имитационной компьютерной модели всей ГС.
Динамика отдельных молекулярно-генетических систем в ОХКММ формально описывается моделями общего вида:
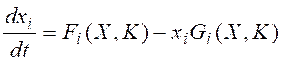
Здесь X = (x1,…,xn) – вектор динамических переменных модели ГС, т.е. концентраций регуляторных продуктов. K = (k1,…,km) – вектор параметров, входящих в уравнения, Fi(X, K) и Gi(X, K) – рациональные функции.
В ОХКММ учитываются временные задержки и локализация процессов в однородных областях, закономерности пассивного и активного переноса веществ и энергии между областями, возможная полиплоидность и гетерозиготность генов, случайные составляющие процессов и пр. В результате изучение динамики функционирования ГС можно рассматривать как математическую проблему анализа решений сложных систем уравнений.
К настоящему времени разработан специальный язык моделирования ОХКММ и программное обеспечение, позволяющее строить имитационные модели сложных многокомпартментных биологических систем, вплоть до организменного уровня.
Проблема адекватности
Одной из наиболее важных и актуальных проблем при построении математических моделей ГС является ее настройка, т.е. побор значений многочисленных констант-параметров, входящих в уравнения. Простейший пример – параметр k в уравнении фосфорилирования на рис. 10.
Как правило, экспериментально измеренные значения известны только для ограниченного числа параметров. Существует также проблема вариабельности набора экспериментальных данных из литературы, по которым оценивают параметры и строят модель. Их получали в конкретных условиях, при разных экспериментальных воздействиях в различные моменты времени и т.д. Недостающие и уточненные данные приходится получать в ходе дополнительных экспериментов с реальными ГС.
Получение значений недостающих параметров назовем настройкой модели. Кроме того, далеко не всегда детально известна структура организации некоторых частей ГС (пути передачи сигналов, механизмы протекания процессов и т.п.). Проверку правильности гипотез о структуре ГС и проверку «устойчивости» ранее полученных оценок параметров при использовании новых экспериментальных данных можно определить как верификацию модели ГС.
Обе проверки можно рассматривать как единую оптимизационную задачу, решение которой проводится на основе специальных численных экспериментов с моделью. Целевой функцией, подлежащей минимизации, выступает общее расхождение реальной экспериментальной динамики от динамики переменных ГС, рассчитанной по ее модели при заданной структуре и параметрах. В качестве динамики могут выступать как зависимости переменных ГС от времени, так и друг от друга.
Рассмотрим, например, метод оптимизации, используемый в разных областях науки и называемый генетическим алгоритмом.
Идея заключается в имитации последовательной «эволюции» популяции, состоящей из так называемых «особей» – моделей генных сетей. Каждая из «особей» обладает «генотипом» – конкретным набором значений параметров модели, несколько отличным от значений у других особей. Допускается также включение в расчет одновременно нескольких моделей сети с несколько отличной структурой. Генотип однозначно определяет «фенотип» – расчетную динамику переменных этой модели генной сети. Особи образуют исходную «популяцию». Она является объектом действия «эволюции», включающей «мутации», «скрещивание родителей» с «рекомбинацией», «размножением потомков», а также «отбор» среди них наиболее приспособленны. Эволюция вызывает изменение состава популяции, которое возникает на каждом эволюционном такте алгоритма. Эволюционный такт, в свою очередь, слагается из двух подтактов.
Первый подтакт состоит в «скрещивании и размножении» особей популяции. Каждая пара моделей-родителей может порождать некоторое количество «потомков». Перед скрещиванием возможен «мутационный процесс», после скрещивания – свободная «рекомбинация» параметров-аллелей. Существует несколько вариантов порождения мутаций. Например, в одном из вариантов от каждого «родителя» возникает несколько «мутантных особей», отличающихся по величине не более чем одного параметра. Небольшие величины отличий задаются методом Монте-Карло случайно для каждой особи. После скрещивания с рекомбинацией возникает новая популяция из родителей и потомков.
Второй подтакт заключается в выделении из нее «наилучшей» субпопуляции. А именно, для каждой особи рассчитывается фенотип (динамика функционирования модели ГС), который сравнивается с едиными и неизменными экспериментальными данными об этой динамике, т.е. вычисляется значение функции приспособленности. Функция приспособленности, подлежащая минимизации, может представлять, например, взвешенное среднеквадратическое расхождение расчетной динамики каждой особи (модели ГС) и «идеала» – экспериментальной динамики. Для последующего размножения отбирается фиксированное количество особей, имеющих наилучшие характеристики индивидуальной «приспособленности». В результате возникает новая популяция, в которой содержатся наилучшие родительские особи и потомки.
После того как новая популяция порождена, такт размножения с мутациями, рекомбинацией и отбором снова повторяется. Постепенно лучшие модели ГС приближаются к реальной экспериментальной динамике.
Только проведя процедуру оптимизации и убедившись в адекватности модели, имеет смысл проводить анализ функционирования ГС на основании расчетов по ее модели.
Поможем написать любую работу на аналогичную тему