При реципрокном направлении скрещивания (самки - дигетерозиготы, самцы - гомозиготны по рецессивным аллелям обоих генов) сочетания признаков исходных родительских линий (Р-поколения), сохранившиеся у потомков анализирующего скрещивания, могут быть нерекомбинантными.
В одном из экспериментов Морган скрещивал черных длиннокрылых самок ![]() с серыми зачаточнокрылыми самцами
с серыми зачаточнокрылыми самцами ![]() . В F1 получено серое длиннокрылое потомство
. В F1 получено серое длиннокрылое потомство ![]() . Потом было произведено анализирующее скрещивание, однако из F1 были взяты не самцы, а самки
. Потом было произведено анализирующее скрещивание, однако из F1 были взяты не самцы, а самки ![]() , которых скрестили с черными зачаточнокрылыми самцами
, которых скрестили с черными зачаточнокрылыми самцами ![]() (рис. 5).
(рис. 5).
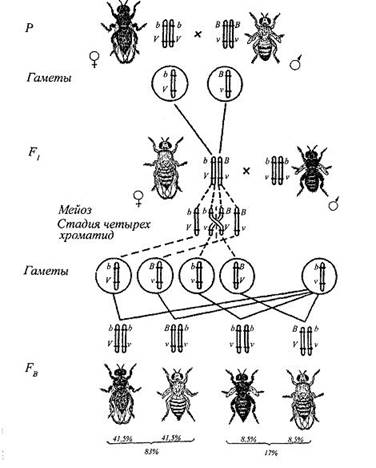
Рис. 5. Кроссинговер у дрозофилы. F2 показан только на самцах: B – серое тело, V – нормальные крылья, v – зачаточные крылья.
При таком скрещивании получено потомство четырех типов: черные длиннокрылые (41,5%), серые с зачаточными крыльями (41,5%), черные с зачаточными крыльями (8,5%) и серые длиннокрылые (8,5%), Таким образом, вместо ожидаемых двух фенотипов, аналогичных родителям, получено два новых сочетания признаков, но с гораздо меньшей частотой. Это значит, что сцепление было неполным.
Полное сцепление генов у дрозофилы наблюдается только в том случае, когда для анализирующего скрещивания берут дигибридных самцов, если же скрещивают дигибридных самок с гомозиготными по рецессивным аллелям самцами, то сцепление никогда не бывает полным.
Причиной неполного сцепления является кроссинговер, который цитоло-гически был открыт Ф. Янссенсом в 1909 г. Изучая мейоз в сперматогенезе у земноводных (саламандру), Янссенс обнаружил, что после конъюгации пары гомологов при расхождении в ряде мест образуются фигуры, имеющие виц греческой буквы хи (χ). Отсюда это явление и получило название хиазмотипии. Т. Морган, опираясь на эти наблюдений Янссенс, высказал гипотезу, что при образовании хиазм гомологичные хромосомы обмениваются участками, несущими блоки генов. Если сцепленные гены лежат в одной хромосоме и у гетерозигот при образовании гамет происходит рекомбинация этих генов, значит, гомологичные хромосомы во время мейоза обменялись своими частями. Обмен гомологичных хромосом своими частями называется перекрестом, или кроссинговером (англ, crossing over означает образование перекреста). Особей с новым сочетанием признаков, образовавшимся в результате кроссинговера, называют кроссоверами.
В результате исследований Морган пришел к выводу, что количество появления новых форм зависит от частоты перекреста, которая определяется по формуле:
Частота перекреста =![]() .
.
Морган установил, что частота перекреста между определенной парой генов - относительно постоянная величина, но различная для разных пар генов. На основании этого сделан вывод о том, что по частоте перекреста можно судить о расстояниях между генами. За единицу измерения перекреста принята его величина, равная 1%, называемая морганидой (сМ). Расстояние между изучаемыми генами влияет на частоту перекреста. Чем дальше расположены гены друг от друга, тем чаще происходит перекрест, и наоборот.
Механизм кроссинговера «разрыв–воссоединение». Согласно теории Янссенса–Дарлингтона, кроссинговер происходит в профазе мейоза. Гомологичные хромосомы с гаплотипами хроматид АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные гаплотипы (хроматиды) Ab и аВ. В анафазе первого деления мейоза происходит расхождение двухроматидных хромосом, а во втором делении – расхождение хроматид (однохроматидных хромосом). Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи. Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи.
Первые прямые цитогенетические доказательства прохождения кроссинговера были получены в 1931 г. Г. Крейтоном и Б.Мак-Клинток на кукурузе и К. Штерном на дрозофиле.
Штерн добился создания гетероморфности для пары XX-хромосом у дрозофилы, используя транслокацию одной хромосомы из пары на XY -хромосому, для другого гомолога транслокацию с четвертой хромосомой (микрохромосомой). В первом случае он получил комплексную хромосому, состоящую почти из всего тела хромосомы и половины Y-хромосомы. Эта хромосома имела необычную форму, отличающую ее от нормальной Х-хромосомы и от других хромосом дрозофилы (рис. 6). Она приобрела Г-образную форму вместо обычной прямой, и в ней был расположен доминирующий ген красных глаз cr+ и рецессивный круглых глаз B+, а вторая часть прикрепилась к маленькой четвертой хромосоме, благодаря чему не терялась при редукционном делении, что было важно для сохранения жизнеспособности таких особей.
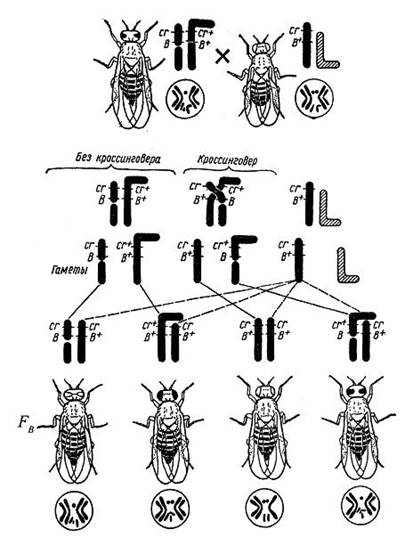 Самки с разными по форме Х-хромосомами были скрещены с самцами, имеющими обычные прямые Х-хромосомы, в которых находились рецессивные гены cr окраски глаз (цвета гвоздики) и круглой их формы В , т.е. самцы были с круглыми глазами гвоздичного цвета. В результате этого скрещивания были получены самки четырех типов: с полосковидными глазами цвета гвоздики
Самки с разными по форме Х-хромосомами были скрещены с самцами, имеющими обычные прямые Х-хромосомы, в которых находились рецессивные гены cr окраски глаз (цвета гвоздики) и круглой их формы В , т.е. самцы были с круглыми глазами гвоздичного цвета. В результате этого скрещивания были получены самки четырех типов: с полосковидными глазами цвета гвоздики ![]() , получившие от матери разделенную на две части Х-хромосому; кругло- и красноглазые
, получившие от матери разделенную на две части Х-хромосому; кругло- и красноглазые ![]() , получившие от матери Г-образную хромосому; в результате кроссинговера - круглоглазые с глазами гвоздичного цвета
, получившие от матери Г-образную хромосому; в результате кроссинговера - круглоглазые с глазами гвоздичного цвета ![]() , у которых должна бьпь короткая изогнутая Х-хромосома с прикрепленным к ней участком Y-хромосомы. и с полосковидными красными глазами
, у которых должна бьпь короткая изогнутая Х-хромосома с прикрепленным к ней участком Y-хромосомы. и с полосковидными красными глазами ![]() . От отца же вес самки должны были получить прямую Х-хромосому.
. От отца же вес самки должны были получить прямую Х-хромосому.
Рис. 6. Цитологическое доказательство кроссинговера у дрозофилы путем параллельного прослеживания хода наследования признаков и поведения при кроссинговере X-хромосом, измененных при помощи двух взаимных транслокаций: cr+ - красная окраска глаз (доминантная), cr – глаза цвета красной гвоздики (рецессивная), B+ - круглые глаза (рецессивный), B – полосковидные глаза (доминантный).
Вследствие кроссинговера происходит рекомбинация – появление новых сочетаний (гаплотипов) наследственных задатков в хромосомах. Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых функционирует как единый суперген, контролирующий несколько признаков. В то же время, в ходе кроссинговера возникают рекомбинации – т.е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
Это означает, что…
а) в ходе естественного отбора в одних хромосомах происходит накопление «полезных» аллелей (и носители таких хромосом получают преимущество в борьбе за существование), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбывают из игры – элиминируются из популяций)
б) в ходе искусственного отбора в одних хромосомах накапливаются аллели хозяйственно-ценных признаков (и носители таких хромосом сохраняются селекционером), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбраковываются).
Поможем написать любую работу на аналогичную тему
Реферат
КРОССИНГОВЕР КАК ПРИЧИНА НАРУШЕНИЯ СЦЕПЛЕНИЯ МЕЖДУ ПРИЗНАКАМИ. КРОССОВЕРНЫЕ И НЕКРОССОВЕРНЫЕ ГАМЕТЫ.
От 250 руб
Контрольная работа
КРОССИНГОВЕР КАК ПРИЧИНА НАРУШЕНИЯ СЦЕПЛЕНИЯ МЕЖДУ ПРИЗНАКАМИ. КРОССОВЕРНЫЕ И НЕКРОССОВЕРНЫЕ ГАМЕТЫ.
От 250 руб
Курсовая работа
КРОССИНГОВЕР КАК ПРИЧИНА НАРУШЕНИЯ СЦЕПЛЕНИЯ МЕЖДУ ПРИЗНАКАМИ. КРОССОВЕРНЫЕ И НЕКРОССОВЕРНЫЕ ГАМЕТЫ.
От 700 руб