По расположению и количеству жгутиков выделяют ряд форм бактерий (рис.1)
1.Монотрихи - имеют один полярный жгутик.
2.Лофотрихи - имеют полярно расположенный пучок жгутиков.
3.Амфитрихи - имеют жгутики по диаметрально противоположным полюсам.
4.Перитрихи - имеют жгутики по всему периметру бактериальной клетки.
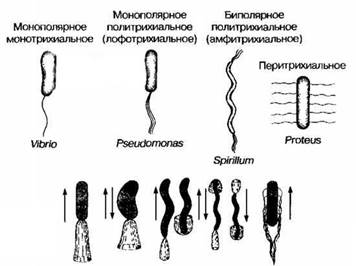
Рис.1. Варианты расположения жгутиков (вверху) и движений бактерий (внизу).
Обычная толщина жгутика — 10 – 20 нм, длина — от 3 до 15 мкм. У некоторых бактерий длина жгутика может на порядок превышать диаметр клетки. Как правило, полярные жгутики более толстые, чем перитрихиальные.
Жгутик состоит из трех частей: нити, крюка и базального тельца (рис.2).
С помощью базального тельца, которое состоит из центрального стержня и колец, жгутик закреплен в цитоплазматической мембране и клеточной стенке. У большинства прокариот нить состоит только из одного типа белка — флагеллина. Аминокислотный состав флагеллина у разных видов бактерий может варьировать. Флагеллин построен из субъединиц с относительно малой молекулярной массой. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
Крюк (толщина 20—45 нм) состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом. Количество колец в базальном теле у грамотрицательных и грамположительных бактерий различно. У грамотрицательных бактерий имеется 4 кольца: L, P, S, M. Из них L и P – наружная пара колец; S и M – внутренняя пара колец. L-кольцо расположено в наружной мембране, P – в пептидогликановом слое клеточной стенки, S – в периплазматическом пространстве, а M – в цитоплазматической мембране.
У грамположительных бактерий базальное тельце устроено проще. Оно состоит только из двух колец: S и M, т.е. только из внутренней пары колец, которые размещаются в цитоплазматической мембране и клеточной стенке.
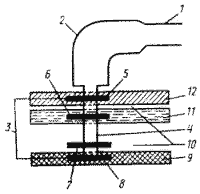
Рис.2. Строение жгутика грамотрицательных эубактерий: 1 — нить; 2 — крюк; 3 — базальное тело; 4 — стержень; 5 — L-кольцо; 6 — P-кольцо; 7 — S-кольцо; 8 — M-кольцо; 9 — ЦПМ; 10 — периплазматическое пространство; 11 — пептидогликановый слой; 12 — наружная мембрана.
Подвижные бактерии могут быть скользящие (передвигаются по твердой поверхности в результате волнообразных сокращений) или плавающие, передвигающиеся за счет движения жгутиков.
Жгутики бактерий по характеру работы подобны корабельному винту. Если клетка имеет много жгутиков, они при движении собираются в пучок, который образует своеобразный пропеллер. Пучок жгутиков, быстро вращаясь против часовой стрелки, создает силу, заставляющую бактерию двигаться почти по прямой линии. После того, как направление вращения жгутиков изменяется, пучок расплетается и бактерия останавливается, вместо поступательного движения она начинает хаотически вращаться, ее ориентация изменяется. В тот момент, когда все жгутики бактерии снова начнут синхронно вращаться против часовой стрелки, образовав пропеллер, толкающий бактерию, направление ее поступательного движения будет отличаться от первоначального. Таким способом бактерия может изменять направление своего движения. Так как у грамположительных бактерий наружная пара колец отсутствует, то считают, что для вращения жгутиков необходимо наличие только внутренней пары (кольца S и M). Эти кольца, соединенные с вращающимся стержнем, выступающим наружу, и образуют так называемый электромотор, обеспечивающий движение жгутика (рис.3). На периферии кольца М находятся белки MotB. Белки MotА встроены в цитоплазматическую мембрану и примыкают к краям колец M и S. Вращающий момент возникает за счет взаимодействия субъединиц белка MotВ с белковыми субъединицами MotА. В белковых субъединицах MotА имеется по два протонных полуканала. Через эти протонные полуканалы переносятся протоны из периплазматического пространства в цитоплазму бактерий (подобно протонному каналу АТФ-синтазы). В результате переноса протонов через белки MotА и MotВ происходит вращение кольца М. Установлено, что один полный оборот кольца М связан с переносом через мембрану около 1000 протонов. Таким образом, в качестве источника энергии для вращения жгутиков используется протондвижущая сила, возникающая в цитоплазматической мембране. Вращение жгутика осуществляется с частотой от 40 до 60 об/с.
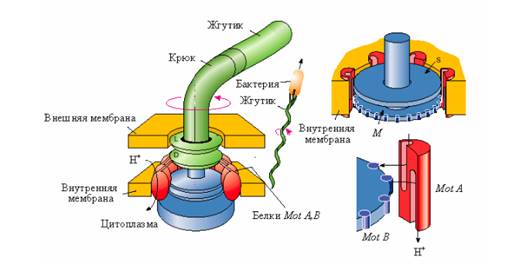
Рис.3. Схематическое изображение электромотора, вращающего жгутики бактерий.
Своеобразный тип движения характерен для спирохет. Клетка спирохет состоит из протоплазматического цилиндра, окруженного цитоплазматической мембраной, и внешнего чехла, представленного пептидогликановым слоем и наружной мембраной. Вокруг протоплазматического цилиндра в периплазматическом пространстве находятся пучки нитчатых структур – аксиальных фибрилл, которые, как и жгутики состоят из белка флагеллина. Эти структуры обеспечивают движение спирохет как в жидкой среде, так и на разделе фаз жидкой и плотной среды (рис.4.).
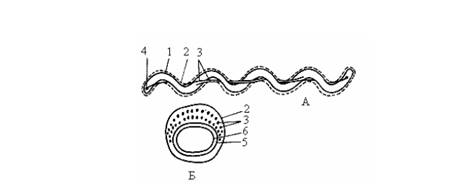
Рис.4. Клетка спирохеты в продольном (А) и поперечном (Б) разрезе.
На рис. А изображена клетка, содержащая по одной аксиальной фибрилле у каждого конца; на рис. Б – поперечный разрез, прошедший через среднюю часть клетки, где показаны два пересекающихся пучка, состоящих из множества аксиальных фибрилл: 1 – протоплазматический цилиндр; 2 – наружный чехол; 3 – аксиальные фибриллы; 4 – место прикрепления аксиальных фибрилл; 5 – пептидогликановый слой клеточной стенки; 6 – цитоплазматическая мембрана.
Число аксиальных фибрилл колеблется от 2 до 100. Один конец каждой оксиальной фибриллы прикреплен вблизи полюса протоплазматического цилиндра, а другой свободный. Клетка содержит по два набора фибрилл, прикрепленных субполярно у каждого клеточного конца. Каждая аксиальная фибрилла тянется почти вдоль всей длины клетки. В центральной части клетки аксиальные фибриллы перекрываются. Фибриллы, вращаясь или сокращаясь, обусловливают характерное для спирохет движение: путем изгибания, вращения вокруг оси, волнообразно, винтообразно.
У некоторых прокариот установлен скользящий тип передвижения. Способность к скольжению выявлена у некоторых микоплазм, миксобактерий, цианобактерий, нитчатых серобактерий и др. Скорость при таком типе передвижения небольшая: 2 – 11 мкм/сек. Общим для всех микроорганизмов, способных к скольжению, является выделение слизи. Кроме того, у ряда таких прокариот в составе клеточной стенки между пептидогликановым слоем и наружной мембраной выявлен тонкий слой белковых молекул. Например, у нитчатых цианобактерий фибриллы образуют единую систему, которая в виде спирали окружает весь трихом (нить). Скольжение нитчатых форм сопровождается и одновременным их вращением, поэтому каждая точка на поверхности трихома описывает при движении спираль. Направление вращения является видоспецифическим признаком и коррелирует с направлением хода спирали белковых фибрилл.
Механизмы скользящего движения не ясны и существуют несколько гипотез, которые их объясняют. Согласно гипотезы реактивного движения скользящее передвижение обусловлено выделением слизи через многочисленные слизевые поры в клеточной стенке, в результате чего клетка отталкивается от субстрата в направлении противоположном направлению выделения слизи. Однако при анализе этой модели было сделано заключение, что для обеспечения скольжения по «реактивному» механизму клетке нужно на протяжении одной секунды выделить такой объем слизи, который во много раз больше ее цитоплазматического содержимого.
В соответствии со второй гипотезой, получившей распространение в последние годы, скользящий тип движения связан с присутствием белкового слоя, который состоит из правильно расположенных фибрилл, аналогичных нитям жгутиков, но находящихся внутри клеточной стенки. У некоторых скользящих бактерий описаны структуры очень схожие с базальными тельцами жгутиковых форм.
Вращательное движение фибрилл, которое «запускается» этими структурами, приводит к появлению на поверхности клетки «бегущей волны» или движущихся микроскопических выпуклостей клеточной стенки, в результате чего клетка отталкивается от твердого или вязкого субстрата. Согласно этой гипотезе, выделение слизи не является абсолютно необходимым для скольжения, но улучшает, в определенных условиях, отталкивание клетки от субстрата. Показано, что скольжение может осуществляться без выделения слизи в среде подходящей консистенции. Более того, выделение большого количества слизи, как правило, мешает передвижению клетки или приводит к его потере.
Для подвижных бактерий характерны таксисы, т.е. направленная двигательная реакция в ответ на определенный раздражитель. По характеру реагирования на раздражение различают положительный таксис — движения по направлению к раздражителю, отрицательные таксисы — движения от раздражителя. В зависимости от действующего раздражителя различают:
1. Хемотаксис – движение бактерий относительно источника химического вещества. Для каждого микроорганизма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы, или эффекторы. Среди эффекторов выделяют: аттрактанты – вещества, которые притягивают бактерии; репелленты – вещества, которые отпугивают бактерии.
2. Фототаксис – движение к источнику света или от него, свойственное фототрофным бактериям.
3. Магнитотаксис – способность бактерий передвигаться по силовым линиям магнитного поля Земли или магнита. Он выявлен в клетках бактерий, содержащих магнитосомы и распространенных в водных экосистемах разного типа.
4. Аэротаксис - передвижение одноклеточных к источнику кислорода или от него.
5. Гидротаксис - движение свободно передвигающихся одноклеточных и колониальных растений и некоторых животных в сторону большей или меньшей влажности.
6. Трофотаксис — активное перемещение в направлении источника пищи
У ряда бактерий выявлен вискозитаксис – способность реагировать на изменение вязкости раствора и передвигаться в направлении ее увеличения или уменьшения. За чувствительность бактерий к градиентам определенных факторов ответственны специфические рецепторы. Изучение хемотаксиса у Escherichia coli позволило обнаружить свыше 30 различных хеморецепторов, представляющих собой белки, синтезируемые независимо от присутствия индуктора или только в результате индукции. Рецептор реагирует на эффектор и передает сигнал по определенному пути, конкретный механизм которого неизвестен, на "мотор" жгутика. У бактерий с перитрихиальным жгутикованием выявлены два вида двигательного поведения: прямолинейное движение и кувырканье, т. е. периодические и случайные изменения направления движения. Если бактерия перемещается в сторону оптимальной концентрации аттрактанта, ее прямолинейное движение, ориентированное по отношению к химическому веществу, становится более длительным, а частота кувырканий более низкой, что позволяет ей в конечном итоге перемещаться в нужном направлении.
Поможем написать любую работу на аналогичную тему