А. Р. Лурия предложил структурно-функциональную модель мозга как субстрата психической деятельности. Эта модель характеризует наиболее общие закономерности работы мозга как единого целого и позволяет объяснить его интегративную функцию (Е. Д. Хомская). Согласно этой модели, весь мозг можно разделить на три структурно-функциональных блока:
а) энергетический блок, б) блок приема, переработки и хранения экстероцептивной информации, в) блок программирования, регуляции и контроля сложных форм деятельности. Любая ВПФ осуществляется при обязательном участии всех трех блоков. Каждый блок характеризуется особенностями строения, физиологическими принципами, лежащими в основе его работы, и той ролью, которую он играет в обеспечении психических функций.
Функциональные блоки мозга. На основе изучения нарушений психических процессов при различных локальных поражениях центральной нервной системы Лурия разработал общую структурно-функциональную модель мозга как субстрата психики. Согласно этой модели весь мозг может быть разделен на три основных блока, характеризующихся определенными особенностями строения и ролью в исполнении психических функций.
1-й блок — энергетический — включает ретикулярную формацию ствола мозга, неспецифические структуры среднего мозга, диэнцефальные отделы, лимбическую систему, медиобазальные отделы коры лобных и височных долей (рис. 16).
Блок регулирует общие изменения активации мозга (тонус мозга, необходимый для выполнения любой психической деятельности, уровень бодрствования) и локальные избирательные активационные изменения, необходимые для осуществления ВПФ. При этом за первый класс активаций несет ответственность преимущественно ретикулярная формация ствола мозга, а за второй — более высоко расположенные отделы — неспецифические образования диэнцефального мозга, а также лимбические и корковые медиобазальные структуры.
Ретикулярная формация (РФ) обнаружена в 1946 г. в результате исследований американского нейрофизиолога Мегоуна, который показал, что эта клеточная функциональная система имеет отношение к регуляции вегетативной и соматической рефлекторной деятельности. Позднее совместными работами с итальянским нейрофизиологом Моруцци было продемонстрировано, что раздражение ретикулярной формации эффективно влияет и на функции
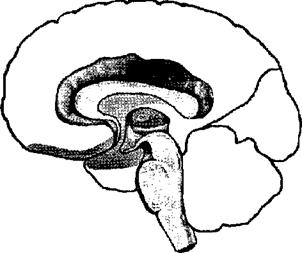
Рис. 16. Функциональные блоки мозга — 1-й блок (по Лурия).
высших структур мозга, в частности коры больших полушарий, определяя ее переход в активное, бодрствующее или в сонное состояние. Исследования показали, что РФ занимает особое место среди других нервных аппаратов, в значительной мере определяя общий уровень их активности. В первые годы после этих открытий было широко распространено представление, что отдельные нейроны РФ тесно связаны друг с другом и образуют однородную структуру, в которой возбуждение распространяется диффузно. Однако позднее выяснилось, что даже близко расположенные клетки РФ могут обладать совершенно различными функциональными характеристиками. РФ расположена на всем протяжении ствола — от промежуточного мозга до верхних шейных спинальных сегментов. Она представляет собой сложное скопление нервных клеток, характеризующихся обширно разветвленным дендритным деревом и длинными аксонами, часть которых имеет нисходящее направление и образует ретикулоспинальные пути, а часть — восходящие. РФ взаимодействует с большим количеством волокон, поступающим в нее из других мозговых структур — коллатералями проходящих через ствол мозга сенсорных восходящих систем и нисходящими путями, идущими из передних отделов мозга (в том числе из двигательных зон). И те и другие вступают с РФ в синаптические связи. Кроме того, многочисленные волокна поступают к нейронам РФ из мозжечка.
Нисходящая часть РФ оказывает неоднозначное влияние на деятельность спинного мозга: раздражение продолговатого мозга (его гигантоклеточного ядра) и некоторых участков варолиева моста сопровождается торможением рефлекторной деятельности нижележащих отделов, а при раздражении более дорсальных и оральных отделов продолговатого мозга — диффузно облегчает действие тех же функциональных структур. Примером первого варианта влияния является эффект мышечного расслабления во время сна. Кроме того, ретикулярное торможение работы нейронов спинного мозга приводит и к ослаблению афферентных восходящих импульсов, то есть снижает передачу сенсорной информации в корковые мозговые центры. Ретикулярные структуры, регулирующие соматические и вегетативные функции, отличаются высокой химической чувствительностью и обнаруживают обратную регулирующую зависимость от характеристик внутренней среды организма (эндокринной системы, уровня СО2в крови и т. п.).
Восходящая часть РФ обеспечивает регуляцию активности высших отделов мозга, главным образом коры больших полушарий. Впервые возможность такого влияния была зарегистрирована в 1935 г. бельгийским нейрофизиологом Бремером в результате перерезки у животных головного мозга на разных уровнях. Поддержание бодрствующего состояния переднего мозга обусловливается первоначальной активацией афферентными раздражениями ретикулярных структур мозгового ствола, а они по восходящим путям определяют функциональное состояние коры, что, конечно, не исключает и прямой передачи афферентации в соответствующие мозговые зоны. Восходящая часть РФ, так же как и нисходящая, помимо деятельности активирующих участков, порождает и общее тормозное влияние. Последнее обеспечивают стволовые участки мозга, в которых найдены так называемые «центры сна», в то время как более дифференцированные по вектору приложения функции предположительно реализуются более высоко расположенными структурами. При исследовании морфологических особенностей клеток РФ было обнаружено, что многие из них имеют Т-образное деление аксонов, один из отростков которых идет вверх, а другой вниз. Это позволило предположить, что и восходящие, и нисходящие функции могут быть связаны с деятельностью одних и тех же нейронов. Кроме того, особенностью отношений коры головного мозга и нижележащих отделов является то, что структуры, обеспечивающие и регулирующие тонус коры, сами находятся с ней в двойных встречных отношениях. Преимущественно тонизируя кору через восходящие пути, лимбические, мезенцефальные и стволовые структуры РФ в то же время подвержены корковой регуляции — и тормозной, и возбуждающей. Эти встречные модулирующие воздействия в первую очередь имеют отношение к лобным отделам, в которых формируются намерения, планы и перспективные программы сознательного поведения. Поведение взрослого человека является примером баланса этих встречных воздействий.
РФ не является спонтанно активирующей системой, а берет энергетический потенциал из двух источников — из обменных процессов организма, лежащих в основе гомеостаза, и из поступающих в организм раздражений внешнего мира. Дефицит во внутренней среде стимулирует инстинктивный компонент поведения, а роль второго источника активности может быть проиллюстрирована эффектом засыпания при искусственном отключении основных рецепторных аппаратов (зрения, слуха и кожной чувствительности).
С точки зрения психических функций энергетический неспецифический блок имеет отношение к процессам общего и селективного внимания, а также к сознанию в целом, процессам неспецифической памяти (залечатлению, хранению и переработке разномодальной информации), к сравнительно элементарным эмоциональным состояниям (страха, боли, удовольствия, гнева). В исполнении последней функции особую роль играют лимбические отделы мозга, которые помимо эмоционального фона обеспечивают переработку интероцептивной информации. Многими учеными РФ рассматривается как водитель многих биологических ритмов организма, часть из которых не только навязывается извне, но и может поддерживаться без видимой внешней стимуляции. Специфическую роль в этой мозговой системе играет не только афферентный энергетический потенциал, но и информационный аспект раздражителя, выражающийся в категориях сенсорного потока и рационального значения (ценности) стимула.
В концентрированной форме специфика работы 1-го блока прослеживается в организации ориентировочного рефлекса: энергетическая мобилизация организма порождается появлением нового стимула, требующего к себе экстренного внимания и сличения с имеющимися в памяти старыми раздражителями, а также последующим переводом полученных итогов в плоскость эмоциональных категорий «вредности-полезности».
2-й блок — приема, переработки и хранения экстероцептивной информации — включает в себя центральные части основных анализаторных систем: зрительной, слуховой и кожно-кинестетической, корковые зоны которых расположены в затылочных, теменных и височных долях мозга (рис. 17). В системы этого блока формально включаются и центральные аппараты вкусовой и обонятельной рецепции, но у человека они настолько оттеснены представительствами высших экстероцептивных анализаторов, что занимают в коре головного мозга незначительное место.
Основу данного блока составляют первичные или проекционные зоны коры (поля), выполняющие узкоспециализированную функцию отражения только стимулов одной модальности. Их задача — идентифицировать стимул по его качеству и сигнальному значению, в отличие от периферического рецептора, который дифференцирует стимул лишь по его физическим или химическим характеристикам. Основная функция первичных полей — тончайшее отражение свойств внешней и внутренней среды на уровне ощущения. Все первичные корковые поля, как это было показано с помощью элек тростимуляции еще канадским нейрохирургом Пенфильдом, характеризуются топическим принципом организации, согласно которому каждому участку рецепторной поверхности соответствует определенный участок в первичной коре («точка в точку»), что и дало основание назвать первичную кору проекционной. Величина зоны представительства того или иного рецепторного участка в первичной коре зависит от функциональной значимости этого участка, а не от его фактического размера.
К числу первичных относятся 17-е (для зрения), 3-е (для кожно-кинестетической чувствительности) и 41-е (для слуха) поля. Вторичные поля представляют собой клеточные структуры, морфологически и функционально как бы надстроенные над проекционными. В них происходит последовательное усложнение процесса переработки информации, чему способствует предварительное прохождение афферентных импульсов через ассоциативные ядра таламуса. Вторичные поля обеспечивают превращение соматотопических импульсов в такую функциональную организацию, которая на уровне психики эквивалентна процессу восприятия. На поверхности мозга вторичные поля граничат с проекционными или окружают их. Номера вторичных полей — 18-е, 19-е, 1-е, 2-е, 42-е, 22-е и частично 5-е. Первичные и вторичные поля относятся к ядерным зонам анализаторов. Третичные (ассоциативные) поля (зона перекрытия) имеют наиболее сложную функциональную нагрузку. Они расположены на границе затылочного, височного и заднецентралъ-ного отделов коры и не имеют непосредственного выхода на периферию. Их функции почти полностью сводятся к интеграции возбуждений, приходящих от вторичных полей всего комплекса анализаторов. Работа этих зон своим психологическим эквивалентом имеет сценоподобное восприятие мира во всей полноте и комбинации пространственных, временных и количественных характеристик внешней среды, но не исчерпывается этим. Второе значение зон перекрытия — это переход от непосредственного наглядного синтеза к уровню символических процессов, благодаря которым становится возможным осуществление речевой и интеллектуальной деятельности. Третичные поля находятся вне ядерных зон. Особого выделения требует зона ТРО (от латинских названий долей: височной — temporalis, теменной — parietalis, затылочной — occipitalis), которая реализует наиболее сложные интегративные функции — 37-е и частично 39-е поле.
Работа второго блока подчиняется трем законам.
Закон иерархического строения. Первичные зоны являются фило- и онтогенетически более ранними. Поэтому недоразвитие первичных полей у ребенка приводит к потере более поздних функций (принцип «снизу-вверх»), а у взрослого с полностью сложившимся психологическим строем третичные зоны управляют работой подчиненных им вторичных и при повреждении последних оказывают на их работу компенсирующее влияние (принцип «сверху-вниз»). Выготский следующим образом характеризует данное теоретическое положение: «Объяснение этой закономерности лежит в том факте, что сложные отношения между различными церебральными системами возникают как продукт развития и что, следовательно, в развитии мозга и в функционировании зрелого мозга должна наблюдаться различная взаимная зависимость центров: низшие центры, служащие в истории развития мозга предпосылками для развития функций высших центров, являющихся вследствие этого зависимыми в своем развитии от низших центров, в силу закона перехода функций вверх сами оказываются в развитом и зрелом мозгу несамостоятельными, подчиненными инстанциями, зависящими в своей деятельности от высших центров. Развитие идет снизу вверх, а распад — сверху вниз».
Закон убывающей специфичности. Наиболее модально специфичными (в данном случае — ориентированными на конкретное свойство объекта, улавливаемое конкретным видом анализатора) являются первичные зоны. Третичные зоны вообще надмодальны.
Закон прогрессирующей литерализации. По мере восхождения от первичных к третичным зонам возрастает дифференцированность функций левого и правого полушария (в основном — по центральным предпосылкам речи и доминантности одной из рук).
3-й блок — программирования, регуляции и контроля за протеканием психической (сознательной) деятельности включает моторные, премоторные и префронтальные отделы коры лобных долей мозга — кпереди от передней центральной извилины (медиобазальные отделы лобных долей входят в первый блок) (рис. 18). Основная цель работы этого блока — формирование планов действий, то есть создание программы психического акта и развертка последовательности исполнения его во времени в реальном поведении. Находясь под постоянным влиянием второго блока, префронтальные (по характеру и сложности обработки информации — ассоциативные) отделы лобных долей одновременно зависимы от речевого и мотивационного компонентов. Подготовка двигательных импульсов завершается их выходом на периферию через двигательную зону коры — преимущественно 4-е поле.
Поможем написать любую работу на аналогичную тему