Для этого необходимо:
- Наличие 2-х раздражителей: безусловного раздражителя и индифферентного (нейтрального) раздражителя, который затем становится условным сигналом;
- Определенная сила раздражителей. Безусловный раздражитель должен быть настолько сильным, чтобы вызывать доминантное возбуждение в центральной нервной системе. Индифферентный раздражитель должен быть привычным, чтобы не вызывать ярко выраженного ориентировочного рефлекса.
- Неоднократное сочетание раздражителей во времени, причем первым должен воздействовать индифферентный раздражитель, затем безусловный раздражитель. В дальнейшем действие 2-х раздражителей продолжается и заканчивается одновременно. Условный рефлекс возникнет в том случае, если индифферентный раздражитель станет условным раздражителем, т.е он сигнализирует о действии безусловного раздражителя.
- Постоянство окружающей среды — выработка условного рефлекса требует постоянства свойств условного сигнала.
Механизм формирования условных рефлексов
При действии индифферентного раздражителя возникает возбуждение в соответствующих рецепторах, и импульсы из них поступают в мозговой отдел анализатора. При воздействии безусловного раздражителя возникает специфическое возбуждение соответствующих рецепторов, и импульсы через подкорковые центры идут в кору головного мозга (корковое представительство центра безусловного рефлекса, которое является доминантным очагом). Таким образом, в коре головного мозга одновременно возникают два очага возбуждения: В коре головного мозга между двумя очагами возбуждения по принципу доминанты, образуется временная рефлекторная связь. При возникновении временной связи изолированное действие условного раздражителя вызывает безусловную реакцию. В соответствии с теорией Павлова, формирование временной рефлекторной связи происходит на уровне коры головного мозга, а в его основе лежит принцип доминанты.
Временная связь (И.П. Павлов - ) - механизм, обеспечивающий функциональную связь между отдельными структурами нервной системы при воздействии двух или более событий актуальной внешней среды и существующая некоторое время. Одним из вариантов проявления временной связи являются условные рефлексы.
Органы чувств устроены так, что они очень живо реагируют на каждый новый раздражитель, который вызывает особый старт-рефлекс, называемый ориентировочным рефлексом, или, как называл его И. П. Павлов, рефлекс "что такое". Например, при звуке звонка уши настораживаются, голова поворачивается к раздражителю. Этот рефлекс помогает животному подготовиться к любым неожиданностям. В коре при этом возникает очаг возбуждения, назовем его А. Если вслед за звуком последует безусловный раздражитель, например появится кормушка с пищей, то в коре возникает второй, более сильный очаг возбуждения Б. Возбуждение центра безусловного рефлекса Б притягивает к себе возбуждение от центра А и между ними замыкается временная связь , образуется новый пищевой условный рефлекс. Новый раздражитель становится сигнальным, или условным, раздражителем, предупреждающим о важном событии, например появлении добычи или хищника. Так образуются пищевые, оборонительные, половые и другие условные рефлексы.
Условные рефлексы первого порядка вырабатываются на базе безусловных рефлексов, т. е. при совпадении во времени действия постороннего, индифферентного раздражителя с какой-либо деятельностью или состоянием организма.
Условные рефлексы могут вырабатываться также на базе прочных условных рефлексов. Например, после того как у собаки образован прочный условным рефлекс на звонок, можно сделать свет условным раздражителем, подкрепляя его не пищей, а звонком. После нескольких сочетаний свет звонок свет, никогда не подкреплявшийся пищей, становится условным пищевым раздражителем. При зажигании лампочки собака "слюнит", поворачивается к кормушке, появляется выраженная пищевая реакция. Такие рефлексы, образованные на базе условных, носят название условных рефлексов второго порядка. У некоторых собак можно с трудом выработать условные рефлексы третьего порядка, у обезьян - до шестого порядка, а у человека надстройка почти неограниченна.
Большое биологическое, а для человека и социальное значение имеют подражательные условные рефлексы. В. Я. Кряжев делил голубей на две группы - "артистов" и "зрителей" - и сажал их в клетку, разделенную сеткой. "Артистам" давался условный сигнал - звонок, который подкреплялся пищей - кормушкой с зерном. Голуби - "зрители" наблюдали. Затем голубей поменяли местами в клетке, и оказалось, что у "зрителей" первое применение звонка вызывало условный рефлекс - движение к кормушке. Таким образом, условный рефлекс необязательно вырабатывается на основе собственного опыта. Он может передаваться через механизм подражания. Об этом всегда должны помнить родители.
Условные рефлексы могут быть выработаны на любой орган, на любую деятельность организма. На почку условный рефлекс вырабатывается следующим путем. Собаке несколько раз через фистулу желудка вливают воду, это вызывает всякий раз повышение диуреза (мочеотделения). Через несколько сочетаний воду, введенную в желудок, тотчас выпускают обратно. Несмотря на это, диурез усиливается. Следовательно, произошло образование условного рефлекса - орошение желудка водой сделалось условным, интероцептивным раздражителем для почки. Примером экстероцептивного рефлекса на внутренние органы - сердце и кровеносные сосуды - может быть следующий опыт. Подкожное введение адреналина вызывает учащение сокращений сердца и сужение сосудов. Если несколько раз звук трубы предшествует введению адреналина, то он сам по себе становится раздражителем, вызывающим те же реакции (учащение сердцебиений, повышение кровяного давления), что и введение адреналина.
Понятие о временной связи. Физиологический механизм, лежащий в основе условного рефлекса, схематически представлен на рис. 1. В зоне коркового представительства условного стимула и коркового (или подкоркового) представительства безусловного стимула формируются два очага возбуждения. Очаг возбуждения, вызванный безусловным стимулом внешней или внутренней среды организма, как более сильный (доминантный) притягивает к себе возбуждение из очага более слабого возбуждения, вызванного условным стимулом. После нескольких повторных предъявлений условного и безусловного раздражителей между этими двумя зонами «проторяется» устойчивый путь движения возбуждения: от очага, вызванного условным стимулом, к очагу, вызванному безусловным стимулом. В результате изолированное предъявление только условного стимула теперь приводит к реакции, вызываемой ранее безусловным стимулом.
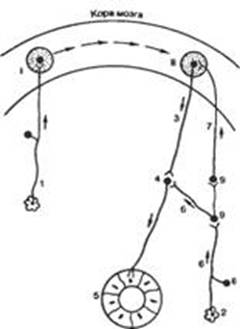 Рис. 1. Образование условного рефлекса (схема). I, II — очаги возбуждения а коре большого мозга; 1 — афферентные пути условного сигнала; 2 — рецепторы полости рта; 3 — эфферентные пути; 4 — центр в продолговатом мозге; 5 — слюнная железа; 6, 7 — афферентные пути безусловного сигнала; 8 — чувствительные нейроны; 9 — вставочные нейроны .
Рис. 1. Образование условного рефлекса (схема). I, II — очаги возбуждения а коре большого мозга; 1 — афферентные пути условного сигнала; 2 — рецепторы полости рта; 3 — эфферентные пути; 4 — центр в продолговатом мозге; 5 — слюнная железа; 6, 7 — афферентные пути безусловного сигнала; 8 — чувствительные нейроны; 9 — вставочные нейроны .
В качестве главных клеточных элементов центрального механизма образования условного рефлекса выступают вставочные и ассоциативные нейроны коры большого мозга.
Павлов пришел к выводу, что замыкание временной связи происходит полностью в коре, между корковым концом анализатора, воспринимающего условный раздражитель, и корковым представительством безусловного рефлекса. После удаления коры условные рефлексы исчезают, а безусловные становятся более грубыми и инертными, менее точными и совершенными, хуже приспособленными к качеству, силе и продолжительности раздражителей. Если удалить у собаки одно полушарие головного мозга, то на поврежденной стороне, несмотря на полную сохранность безусловно-оборонительного рефлекса, не может быть выработан не его базе условный рефлекс, тогда как на здоровой стороне он легко вырабатывается. .
|
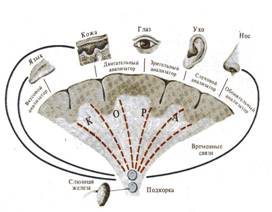
Рис. 2. Формирование условного рефлекса. Первоначальная схема И. П. Павлова . Внимание!
Если вам нужна помощь в написании работы, то рекомендуем обратиться к
профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные
корректировки и доработки. Узнайте стоимость своей работы.
|
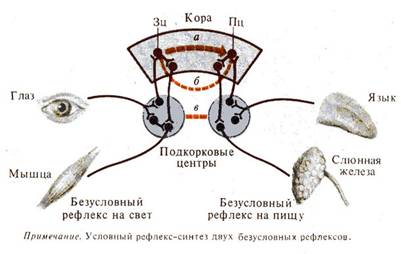
Рис. 3. Формирование условного рефлекса. Схема Э. Асратяна (1956): а, б, в — уровни замыкания; красный пунктир — временная связь |
Последующие нейрофизиологические исследования привели к разработке, экспериментальному и теоретическому обоснованию других гипотез об образовании условного рефлекса (рис. 3). Данные современной нейрофизиологии указывают на возможность разных уровней замыкания, формирования условно-рефлекторной связи (кора — кора, кора — подкорковые образования, подкорковые образования — подкорковые образования) при доминирующей роли в этом процессе корковых структур. Очевидно, физиологический механизм образования условного рефлекса представляет собой сложную динамическую организацию корковых и подкорковых структур мозга (Л. Г. Воронин, Э. А. Асратян, П. К. Анохин, А. Б. Коган). Дальнейшие исследования структуры дуги безусловного рефлекса показали что она в своей центральной части состоит из многих параллельных ветвей, проходящих через различные уровни ЦНС.
Корковый пункт условного раздражителя в принципе имеет такую же структуру, как корковое представительство безусловного рефлекса, так как условный раздражитель сам по себе вызывает соответствующую безусловно-рефлекторную реакцию, особенно, если в качестве условного раздражителя взять какой-нибудь механический, электрический или химический раздражитель. Поэтому при сочетании условного и безусловного раздражителей временная связь по существу замыкается между корковыми представительствами двух безусловных рефлексов, т.е. между нейронами корковых ветвей их рефлекторных дуг. Следовательно, процесс образования условного рефлекса можно рассматривать как корковый синтез двух (или нескольких) безусловных рефлексов.
Такое представление о месте и характере замыкания временной связи подтверждается экспериментальными исследованиями Э.А. Асратяна. Если подобрать два безусловных раздражителя примерно одинаково физиологической силы, например, пищевой и оборонительный, а затем сочетать их строго попеременно (то в одной, то в обратной последовательности), то образуется условный рефлекс в необычной форме. Каждый из безусловных раздражителей будет вызывать как двигательную, так и секреторную реакцию. Электрокожное раздражение становится сигналом пищи, а пища - сигналом электрокожного раздражения. Следовательно‑ временные связи могут быть двусторонними.
При исследовании роли РФ было установлено, что разрез по всей толще коры полушарий, разобщающий друг от друга центры условного и безусловного раздражителей, не препятствуют осуществлению ранее выработанного рефлекса или образованию нового. Эти факты послужили основанием для утверждения, что замыкание временной связи может осуществляться в РФ по схеме: кора-подкорка-кора.
Однако приведенный факт не может служить еще бесспорным и прямым доказательством того, что временные связи замыкаются не в коре. Так, показано, например, что двигательный пищевой условный рефлекс на свет сохраняется у животного после операции "подрезки" участков зрительной и двигательной зон коры, т.е. когда пересекались пути, восходящие к этим участкам от РФ. По-видимому, более верным будет считать, что временные связи могут образовываться на различных уровнях ЦНС, и, как показано в последнее время, даже на уровне вегетативных ганглиев, однако инициативная и ведущая роль в этом процессе в естественных условиях принадлежит корковым клеткам. На основании морфологических и электрофизиологических исследований, в частности, электрической активности клеток различных слоев коры во время образования УР высказывается предположение, что временная связь замыкается, вероятно, на клеточных телах вставочных нейронов. Сам процесс замыкания временной связи заключается, по мнению многих ученых, в том, что ранее бездействующие синапсы вставочных нейронов становятся проходимыми для нервных импульсов при сочетании условного и безусловного раздражителей.
Согласно гипотезе конвергентного замыкания условного рефлекса П.К. Анохина индифферентный и безусловный раздражители вызывают генерализованную активацию коры (благодаря участию РФ) т.е. каждый из них вызывает сразу возбуждение разнообразных нейронов в различных областях коры. При сочетании этих раздражителей имеет место конвергенция двух восходящих возбуждений (индифферентного и безусловного) по клеткам коры больших полушарий. Происходит перекрытие этих возбуждений на одном и том же корковом нейроне и их взаимодействие. В результате такого взаимодействия возникают и стабилизируются временные связи на различных корковых элементах, находящихся в различных участках мозга.
Физиологические механизмы замыкания временной связи. Образование временной связи, по Павлову, является результатом взаимодействия двух одновременно возбуждаемых пунктов коры. Наличие двух очагов возбуждения в коре закономерно вызовет движение процесса возбуждения от более слабого (вызванного индифферентным раздражителем) к более сильному ( вызванному безусловным раздражителем). Таким образом, в основе механизма замыкания временной связи Павлов усматривал явление типа проторения пути, суммационного рефлекса, доминанты.
Сохранив принцип конвергенции стимулов гетерогенных модальностей как исходный механизм взаимодействия и интеграции, П.К. Анохин (1968, 1974) дополнил концепцию представление о том, что взаимодействие сигналов различных сенсорных и биологических модальностей происходит на одном нейроне (рис. 4). Особые свойства подкрепляющего фактора определяются специфическими химическими реакциями, которые он вызывает, и соответствующими молекулярными преобразованиями в пределах мембраны и цитоплазмы нейрона. Здесь происходит ассоциация эффектов условной и безусловной стимуляции с последующими выходами сложившейся интеграции на аксон в виде клеточного разряда. Данная теоретическая концепция является основой для проведения исследований механизмов формирования ассоциативного обучения на одном нейроне.
Микроэлектродные исследования показали, что формирование ассоциативных связей происходит на всех уровнях мозга млекопитающих. Исследованы нейроны коры, ретикулярной формации, лимбических структур, стриопаллидарной системы, гипоталамуса. Показано, что до 60% нейронов различных корковых и подкорковых структур мозга способны формировать условные ответы на ранее неэффективные стимулы.
Внутриклеточные механизмы ассоциативного обучения. Опыты с ассоциативным обучением изолированных нейронов показали, что около 15% из них обучаются в процессе предъявления сочетаний. и около 80% - демонстрировали феномен отсроченного обучения, когда реакция на условный сигнал появлялась не сразу, а через какое-то время. Независимо от вида ассоциируемых стимулов и от особенностей предъявления сочетаний, каждый конкретный нейрон всегда обучается по одному и тому же способу – или во время обучения, или отсроченно. Это качество является его индивидуальной характеристикой. В основе этого феномена лежат особенности внутриклеточных процессов, опосредующих ассоциативное обучение, и эти процессы различны по скорости их развития.
Эффективность входа в нейрон может меняться под влиянием разной степени эффективности последующих возбуждений, поступающих по другим входам в этот же нейрон. С этой точки зрения особый интерес представляют командные нейроны, рецептивное поле которых состоит из широкой пластичной зоны и узкой области высоко стабильных реакций. При повторении стимула на пластичную зону рецептивного поля реакция угасает. При нанесении стимула на зону стабильных реакций ответы нейрона устойчиво сохраняются. При комбинации раздражений малоэффективной зоны с раздражениями области эффекторных реакций ответы со слабо эффективной зоны усиливаются. В пределах одного нейрона неэффективный вход может стать эффективным при сочетании его возбуждения с возбуждением высокоэффективного входа. С этой точки зрения формирование временной связи возможно в отдельном нейроне.
Поможем написать любую работу на аналогичную тему