1. В настоящее время известны три разных механизма темновых реакций фотосинтеза у высших растений. Но, по-видимому, правильнее говорить об одном основном процессе и двух вариантах.
Основной механизм – это фиксация углерода в цикле Кальвина. В последнее время этот цикл стали называть С3-путем, или С3-типом, фотосинтеза, а растения, осуществляющие только реакции этого цикла, называют С3-растениями. Такие растения обычно растут в областях умеренного климата; оптимальная дневная температура для фиксации углекислого газа у этих растений составляет от +15 до +25 °С.
Первый вариант – это С4-путь (или С4-тип фотосинтеза), называемый также циклом Хетча–Слэка. Растения, осуществляющие данный тип фотосинтеза, распространены в тропических и субтропических областях.
Второй вариант – процесс, известный под названием метаболизма органических кислот по типу толстянковых (МОКТ- или САМ-фотосинтез). Растения с таким типом фотосинтеза часто встречаются в засушливых пустынных областях.
Весь цикл фиксации углерода можно представить в виде следующих стадий.
ü Первая стадия – непосредственная фиксация углекислого газа – карбоксилирование.
ü Вторая стадия – образование 3-фосфоглицеринового альдегида (ФГА).
ü Третья стадия – образование продуктов фотосинтеза.
ü Четвертая стадия – восстановление первоначальных реагентов.
Перечисленные стадии выделены условно – вместе они составляют цикл фиксации углерода, или цикл Кальвина.
Карбоксилирование
Эта стадия – ключевая, потому что в ней участвует СО2. Молекула углекислого газа соединяется с молекулой пятиуглеродного сахара рибулезодифосфата (РДФ) с образованием нестойкого шестиуглеродного соединения, которое затем распадается на две молекулы 3-фосфоглицериновой кислоты
Реакция карбоксилирования очень интересна тем, что в зависимости от условий она может протекать с образованием различных конечных продуктов. Так, например, при наличии СО2 продуктом реакции будет только ФГК, а в присутствии О2 РДФ не присоединяет углекислый газ и распадается на ФГК и фосфогликолевую кислоту, которая используется в процессах фотодыхания. Фотодыхание – это процесс, протекающий лишь на свету и сопровождающийся поглощением О2 и выделением СО2. Такое изменение хода реакции объясняется тем, что участвующий в ней фермент обладает двойной каталитической активностью – по отношению к углегислому газу и кислороду.
Этот фермент называется рибулозо-1,5-бифосфаткарбоксилаза-оксигеназа (РуБФ-карбоксилаза). Этот фермент составляет около 50% всех растворимых белков в листьях и потому может считаться самым распространенным белком в природе. Фермент состоит из двух субъединиц – большой и малой. Интересно, что белки больших субъединиц кодируются ДНК хлоропластов, а белки малых субъединиц – ядерной ДНК. Большие субъединцы обладают каталитической активностью и в отсутствие малых, которые, по-видимому, играют регуляторную роль. Этот факт может служить подтверждением того, что хлоропласты произошли от прокариотических предков.
Таким образом, на первых этапах фиксации углерода имеет место конкуренция между двумя процессами – фиксацией углерода и фотодыханием. Для сдвига баланса в сторону фиксации углерода необходимы ионы Мg2+ (2).
Образование фосфоглицеринового альдегида
Образующаяся на первой стадии ФГК превращается в ФГА в два этапа (3 и 4). Сначала используется АТФ, синтезированная в световой фазе фотосинтеза. Затем используется НАДФ.Н, который тоже является продуктом световой фазы фотосинтеза.
Молекула ФГА является ключевым веществом для третьей стадии.
Образование продуктов фотосинтеза
Обычно продуктом фотосинтеза называют сахар. На самом деле продуктами фотосинтеза можно считать и другие вещества, о чем мы упоминали при рассмотрениии Z-схемы.
Молекула ФГА используется растением в цикле Кальвина в нескольких направлениях.
- Во-первых, ФГА является основой для синтеза сахара.
- Во-вторых, ФГА может быть использована для синтеза аминокислот.
Среди продуктов фотосинтеза обнаружены такие аминокислоты, как аланин, серин, глютаминовая кислота, глицин. Синтез аминокислот происходит интенсивно при недостатке НАДФ.Н, в результате чего из ФГК образуется не ФГА, а пировиноградная кислота, которая является исходным соединением для синтеза аминокислот и одним из ключевых веществ цикла Кребса. - В-третьих, ФГА дает начало циклу превращений некоторых промежуточных продуктов в РДФ, который служит акцептором углекислого газа.
Наряду с углеводами и аминокислотами из промежуточных продуктов цикла Кальвина могут образовываться липиды и другие продукты.
Во всех уравнениях фотосинтеза в правой части пишется формула шестиуглеродного сахара. Как правило, его называют глюкозой. Но в действительности первым свободным сахаром является дисахарид сахароза, из которой образуются два моносахарида – глюкоза и фруктоза.
Восстановление первоначальных реагентов
Для того чтобы растение могло акцептировать новую молекулу углекислого газа, необходимо иметь РДФ, основной акцептор углекислого газа. РДФ образуется из ФГА в результате цепи реакций, в процессе которых образуются пяти- и семиуглеродные сахара. Надо отметить, что основная масса ФГА идет именно на восстановление нужного количества РДФ: из 12 образовавшихся молекул ФГА только две идут на образование продуктов фотосинтеза, т.е. сахарозы.
Подводя итог рассмотрению фаз фотосинтеза, можно составить обобщенную схему фотосинтеза (рис. 1).
Фиксация углекислого газа у С3- и С4-растений значительно различается. Если у С3-растений молекула углекислого газа присоединялась к пятиуглеродной молекуле РДФ, то у С4-растений акцептором углекислого газа является трехуглеродная молекула, чаще всего – это фосфоенолпировиноградная кислота (ФЕП). Соединяясь с углекислым газом ФЕП превращается в щавелевоуксусную кислоту (ЩУК), которая и поступает в хлоропласт клеток мезофилла. В хлоропластах ЩУК при наличии НАДФ.Н превращается в яблочную кислоту (ЯК), которая поступает в клетки обкладки сосудистых пучков. В клетках обкладки сосудистых пучков ЯК отдает молекулу углекислого газа в цикл Кальвина, превращаясь в пировиноградную кислоту (ПВК). ПВК, в свою очередь, возвращается в хлоропласты мезофилла, превращается в ФЕП, и начинается новый цикл (рис. 3).
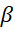
Увеличение числа реакций для фиксации углекислого газа у С4-растений на первый взгляд может показаться излишним и бессмысленным. Но это только на первый взгляд. Растениям с С4-типом фотосинтеза приходится концентрировать углекислый газ в клетках обкладки, т.к. по сравнению с С3-растениями в их клетках углекислого газа содержится значительно меньше. Это связано с тем, что С4-растения обитают в более жарком и сухом климате, чем С3-растения, поэтому для уменьшения потерь воды им приходится уменьшать транспирацию. За счет этого создаются трудности в поглощении углекислого газа, что и приводит к необходимости его концентрации. В настоящее время считается, что С4-тип фотосинтеза является эволюционным приспособлением к более жарким и сухим климатическим условиям.
Поможем написать любую работу на аналогичную тему