Конструирование молекулярных векторов невозможно без набора ферментов, которые можно назвать молекулярными инструментами генетической инженерии. Основные процедуры, которые проводят с ДНК при создании векторов, это разрезание и сшивка молекул ДНК. Соответственно, основными инструментами являются ферменты эндонуклеазы (рестриктазы) и лигазы.
Также применяются ферменты для достройки комплиментарной цепи – полимеразы, гидролиза одноцепочечных концов – экзонуклеазы, создании комплиментарной ДНК на матрице РНК – обратная транскриптаза, присоединяющие дезоксинуклеотиды к одноцепочечным концам ДНК – трансферазы.
Таким образом, при манипуляции с векторами полезны все ферменты, субстратом для которых являются нуклеиновые кислоты.
Эндодезоксирибонуклеазы
(= эндонуклеазы, эндонуклеазы рестрикции, сайтспецифические энодезоксирибонуклеазы, рестриктазы, restriction enzyme or restriction endonuclease)
Открытие рестриктаз произошло в ходе изучения в 50-х годах развития бактериофага λ на бактерии Escherichia coli. Было обнаружено, что 2 штаммы E. coli, на которых размножались фаги, производили с ними некую модификацию, после которой фаг, полученный на одном штамме, ограниченно развивался на другом штамме. Установили, что это связано с разрушение ДНК фага с помощью разрезающих ферментов, и одновременно модификацией (метилированием; methylation, добавление метильной группировки -CH3) части молекул, защиты их от разрезания, с помощью другой группы ферментов – ДНК-метилаз (ДНК-метилтрансфераз). Таким образом, немногочисленные фаги с метилированной ДНК выживали и размножались, но метилирование не наследовалось. ДНК хозяина защищена метилированием.
Термин «рестриктазы» происходит от слова «ограничение» – т.е. такое название дано от явления ограниченного развития фагов на определенных штаммах бактерий.
Субстрат рестриктаз – двунитевая молекула ДНК. В основе расщепления лежит реакция гидролиза. Таким образом, в бактериальной клетке иммееся два типа ферментативной активности: расщепление рестриктазой по сайту узнавания и защитное метилирование сайта узнавания (R-M-система). Последнее (M) защищает собственную ДНК от разрушения. Чужеродная же ДНК, попадающая в клетку, служит субстратом для обоих ферментов.
Как было установлено позднее, основное назначение рестриктаз – ограничение скрещиваний между видами бактерий и иногда штаммами бактерий. Тем не менее, не все скрещивания ограничиваются, и обмен генетическим материалом все-же происходит и дает эволюционное преимущество рекомбинированной клетке. R-M-система только сдерживает внедрение чужеродного генетического материала, поддерживая, таким образом, генетическую обособленность видов.
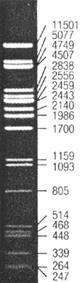
Первая рестриктаза была выделена Мезельсон и Юань в 1968 г. и называлась EcoK. Однако этот фермент расщеплял ДНК не по сайту узнавания, а рядом с ним. Поэтому поиск рестриктаз пошел по пути выделения тех, которые специфически узнают и сцецифически расщепляют определенные локусы ДНК. Таким образом, при расщеплении чистой линии фага λ, или других наборов идентичных молекул ДНК, получается набор фрагментов строго определенного размера, который часто используют для сравнительной оценки размера молекул при электрофоретическом разделении (иллюстрация ниже).
|
|
λ / PstI - размерный молекулярный маркер для ДНК. Представляет собой геном чистой линии фага лямбда, разрезанный рестриктазой PstI. Слева от электрофоретического профиля даны размеры фрагментов в парах оснований. |
Номенклатура данных ферментов была предложена в 1973 г. Smith & Natans. Название рестриктазы включает следующие пункты.
(1) трехбуквенное название, производное от бинарного названия микроорганизма-источника: Eco – Escherichia coli, Hin – Haemophilus influenzae, Pst – Providentia stuartii, Sal – Streptomyces albus.
В случае «похожести» видовых названий акронимы могут строиться по несколько другому принципу: Haemophilus parainfluenzae – Hpa, Haemophilus parahaemolyticus – Hph.
(2) в случае необходимости, обозначение серотипа или штамма вслед за акронимом родо-видового названия: Hind – Haemophilus influenzae d, Escherichia coli B – EcoB.
(3) различные R-M системы, кодируемые одной клеткой, обозначаются латинскими цифрами: HindI, HindII, HindIII.
(4) если система генетически локализована в геноме фага или на плазмиде, то после родо-видового названия указывается символ внехромосомного элемента: EcoPI, EcoRII. Штаммовая принадлежность может указываться в скобках: Eco(K)PI.
(5) Иногда после родо-видового акронима указывают номер бактериальной культуры в коллекции: например, Pvu84II, Hinb1076III.
Молекулярная структура: Рестриктазы класса I состоят из субъединиц, или доменов, α, β и γ, обладающих различной молекулярной массой. Субъединицы обладают разными функциями: узнавание последовательности (γ), расщепление ДНК (α), метилирование ДНК (β). Рестриктазы класса II состоят из 2 субъединиц.
Классификация большого (на сегодняшний день) количества известных рестриктаз основана в основном на характере расщепления ДНК.
|
потребность в кофакторах |
характер расщепления |
|
|
Класс I (системы R-M класса I) |
аденозинтрифосфат (ATP), S-аденозинмонофосфат (SAM), ионы Mg2+; расщепление ДНК совмещено с гидролизом АТФ |
случайным образом на расстоянии 400-7000 п.н. от сайта узнавания, продукты расщепления гетерогенны (модель работы: два фермента после узнавания двух сайтов на одной молекуле ДНК начинают перемещать – транслоцировать – цепь ДНК через себя, пока не встретятся, и тогда в точке встречи происходит разрезание) |
|
Класс II (системы R-M класса II) – ферменты R и M обособлены |
ионы Mg2+ |
рестриктазы выделяются свободными от метилазной активности; узнавание и разрезание по одному и тому же сайту; сайт узнавания включает 4–8 п.н., и иметь ось симметрии 2-го порядка (является палиндромом); a) расщепляющие по оси симметрии с образованием тупых концов; б) расщепляющие с образованием 5’-липких концов; липкие концы одинаковы, и поэтому фрагменты в смеси могут слипаться; возможна стыковка фрагментов ДНК из любых неродственных организмов; в) расщепляющие с образованием 3’-липких концов |
|
Класс II, подкласс IIS (S от слова «shift») |
Расщепляют ДНК на строго фиксированном расстоянии от участка узнавания |
|
|
Класс II, подкласс искусственных рестриктаз* - являются продуктами белковой инженерии |
||
|
Класс III |
АТФ, ионы Mg2+
|
Обладают рестриктазной и метилазной активностью; узнают несимметричные последовательности длиной 5-6 п.н. и расщеплют цепь на расстоянии 24-27 п.н. от участка узнавания, образую одноцепочечные 5´-концы дл. 2-3 нуклеотида |
*Artificial restriction enzymes can be generated by fusing a natural or engineered DNA binding domain to a nuclease domain (often the cleavage domain of the type IIS restriction enzyme FokI). Such artificial restriction enzymes can target large DNA sites (up to 36 bp) and can be engineered to bind to desired DNA sequences.
Для характеристики рестриктазы схематически показывают узнаваемую последовательность и место расщепления, причем достаточно показать схему только на одной цепи; комплиментарная цепь расщепляется симметрично. Например:
PstI: CTGCA ↓ G
G↑ ACGTC
Ряд рестриктаз класса II способны расщеплять несколько разных последовательностей с общей формулой (частично вырожденные последовательности):
HindII: GTPy ↓ PuAC
EcoRII: ↓ CC(AT)GG
HaeI: (AT)GG ↓ CC(TA)
Соответственно в продуктах одной рестрикции образуются разные типы липких концов, и не все полученные фрагменты могут в дальнейшем быть объединены между собой.
Размер узнаваемой последовательности связан с частотой встречаемости данных участков в природной ДНК. Соответственно, длинные последовательности узнавания встречаются реже.
Ферменты, узнающие послед. из ≥6 п.н. называют крупнощепящими (редкощепящими), а 4-х п.н. – мелкощепящими (частощепящими).
Особый интерес представляют рестриктазы, дающие комплиментарные липкие концы.
Так, если соединить липкие концы, образованные BamHI и BglII, то после их объединения образуется последовательность, не узнаваемая ни BamHI, ни BglII, но узнаваемая Sau3AI:
5’…NNG 3’ 5’GATCTNN…3’
3’…NNCCTAG 5’ + 3’ANN…5’
↓ ДНК-лигаза
5'…NNGGATCTNN…3’
3’…NNCCTAGANN…5’
В процессе открытия новых рестриктаз их называют собственно новые, если они выделены из ранее неизученного штамма. Рестриктазы делят на прототипы – новые рестриктазы, узнающие совершенно новую последовательность и изошизомеры – узнающие одинаковые последовательности. Часто новые рестриктазы узнают последовательности, для которых уже известны прототипы. Иными словами, количество различных последовательностей узнавания равно количеству прототипов. К 2002 году было открыто 211 прототипов.
Истинные изошизомеры – кроме того, это те изошизомеры которые одинаково расщепляют цепи. В ином случае, если последовательности узнавания одинаковы, а способы расщепления различны – это будут ложные изошизомеры.
Кроме того, рестриктазы делятся на две группы по способности расщеплять (а) немодифицированную и (б) метилированную ДНК (например, DpnI).
Было установлено также, что частота расщепления ДНК по данной последовательности узнавания зависит от нуклеотидного состава участков, примыкающей к ней, и может отличаться в 4 раза. По этой причине при конкретных манипуляциях с ДНК для исчерпывающего расщепления берут некоторый избыток фермента.
В то же время, такие факторы, как избыток фермента, снижение ионной силы раствора, повышение pH, наличие органических растворителей в смеси приводят к неспецифическому действию рестриктаз. Например, для EcoRI неспецифичность выражается в узнавании укороченной последовательности ↓AATT вместо G↓AATTC.
Точность гидролиза рестриктазами in vivo повышается за счет присутствия в прокариотических клетках особого вещества, спермидина, относящегося к полиаминам.
Наиболее интересны для целей молекулярного клонирования рестриктазы образующие достаточно длинные «липкие концы».
Направленное введение разрыва с помощью рестриктаз IIS производится с помощью олигонуклеотида, одна часть которого комплиментарна участку на одноцепочечной ДНК, который необходимо разрезать, а вторая часть образует двухцепочечную шпильку с последовательностью узнавания рестриктазой IIS. Примеры рестриктаз IIS: BinI, FokI, TaqII.
Поможем написать любую работу на аналогичную тему