Уже подчеркивалось, что чужДНК интегрируется в клеточные хромосомы в случайных локусах при любом способе ее введения. При работе с трансгенными животными, полученными с помощью вышеописанных методов, это означает вынужденную констатацию такого факта и его последующий анализ. Действительно, в данном случае провести предварительный анализ результатов интеграции чужДНК на уровне зародышевых клеток невозможно, поскольку необходим этап культивирования. Положение спасают различные культивируемые линии эмбриональных стволовых клеток. Уже говорилось, что они являются плюрипотентными и у взрослых животных дают начало многим типам клеток, включая зародышевые. Таким образом, можно трансформировать эти клетки рекДНК или инфицировать рекомбинантным ретровирусом, отобрать желаемую линию, а потом перенести рекомбинантное ядро с помощью микропипетки в оплодотворенную яйцеклетку, из которой затем удаляют оба пронуклеуса. Полученное потомство будет трансгенным. При необходимости, перенося ядра генотипически одинаковых плюрипотентных клеток, можно создавать клоны трансгенных животных.
Перенос рекомбинантных стволовых клеток в бластоцисты и последующая имплантация последних "приемным" матерям приводят к появлению химерных потомков (рис. 1.3.1 а, б, в). У взрослых животных доля тканей, происходящих из внесенных стволовых клеток, варьирует в широких пределах от эксперимента к эксперименту; такой тканью может быть и зародышевая. Для выявления этого случая животных скрещивают с особью нормального генотипа и анализируют потомство на появление в нем рекомбинантных признаков (рис. 1.3.1, г, д). Такие особи будут трансгенными гетерозиготами.
Эмбриональные стволовые клетки используют также для получения трансгенных животных, у которых произведено замещение какого-либо гена на его аллель. Для исследования свойств мутантных аллелей, в том числе нуль-аллелей, крайне важно, чтобы они находились в обычных для них локусах. Естественным методом замещения является гомологическая рекомбинация, но у животных этот процесс происходит крайне редко. При трансформации клеток гомологичной ДНК она преимущественно интегрируется в случайных локусах, и только небольшая часть внедрений (10-3—10-2) происходит благодаря рекомбинации через гомологичный сайт. Частота гомологической рекомбинации пропорциональна размеру взаимодействующих участков и достигает насыщения (100-кратного увеличения частоты) при длине около 14 т.п.н., причем рекомбинация происходит главным образом в ранней S-фазе .
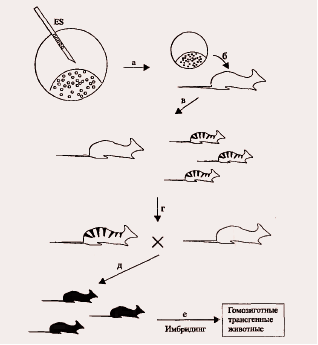
Рис. 1.3.1. Метод получения линии трансгенных мышей с использованием эмбриональных стволовых клеток.
Поможем написать любую работу на аналогичную тему