Фотосинтез бактериальный— фотосинтез, осуществляемый бактериальными микроорганизмами. В отличие от растений, фотосинтез у пурпурных и зеленых бактерий не сопровождается выделением кислорода, так как донорами водорода при ассимиляции углекислоты в этом процессе служит не вода, а другие вещества: сероводород, молекулярная сера, тиосульфат, сульфит, молекулярный водород, органические соединения. Соответственно природе водородного донора образуются различные окисленные продукты. Источником углерода при Ф. б. может быть не только углекислота, но и некоторые органические соединения, в частности, ацетаты, пируваты. У синезеленых водорослей (цианобактерий), как и у растений, в процессе Ф. б. в качестве донора водорода образуется вода и выделяется кислород.
У прокариот обнаружены следующие типы фотосинтеза:
– зависимый от бактериохлорофиллов бескислородный фотосинтез;
– зависимый от хлорофиллов кислородный фотосинтез;
–зависимый от бактериородопсина бескислородный фотосинтез галофильных бактерий.
Структурная организация фотосинтетического аппарата прокариот представлена тремя компонентами:
1. Светособирающие пигменты поглощают свет и передают в реакционные центры.
2. Фотохимические реакционные центры трансформируют энергию.
3. Фотосинтетические электронтранспортные системы обеспечивают перенос электронов, сопряженный с запасанием энергии в АТФ.
Современные фотоавтотрофы используют свет видимой и ближней инфракрасной части спектра. Они содержат магний-порфириновые пигменты, набор которых характерен и постоянен для определенных групп.
Пигменты относятся к двум классам соединений: 1) пигменты с тетрапиррольной структурой (хлорофиллы, фикобилипротеиды); 2) пигменты, основу которых составляют полиизопреноидные цепи (каротиноиды). Способность улавливать свет зависит от системы двойных сопряженных связей, то есть систем с чередующимися двойными и простыми связями.
Известно около 10 видов хлорофиллов. Хлорофилл а обеспечивает поглощение света до 700–740 нм, встречается у цианобактерий, у всех водорослей и у высших растений. Бактериохлорофилл а имеет максимумы поглощения в диапазоне длин волн от 375 до 890 нм. Этот пигмент содержат большинство бактерий – фотосинтетиков. Бактериохлорофилл в найден у пурпурных бактерий, поглощает свет в длине волны до 1100 нм, максимум поглщения приходится на 1020–1040 нм. Бактериохлорофиллы с и d позволяют зеленым серобактериям использовать свет с длиной волны до 840 нм. Гелиобактерии также содержат бактериохлорофилл d.
Фикобилипротеиды – красные и синие пигменты цианобактерий. Хромофорная структура пигментов – фикобилин, состоит из четырех пиррольных колец, развернутых и не содержащих металла. Обнаружены три типа: С-фикоцианин, С-фикоэритрин, С-аллофикоцианин. Фикобилипротеиды обеспечивают поглощение света в области 450–700 нм, а также высокоэффективную передачу (до 90%) поглощенного света на хлорофилл.
Каротиноиды-вспомогательные пигменты, продукты конденсации изопрена. У алифатических каротиноидов цепь развернута, у моно- и бициклических каротиноидов на концах цепи расположены ароматические кольца. Кислородсодержащие каротиноиды относят к группе ксантофиллов. Каротиноиды поглощают свет в синем и зеленом участке спектра, в длине волны 400–550 нм. Выделено свыше 20 каротиноидов пурпурных бактерий, наиболее распространены каротины, ксантофиллы, определяющие окраску от желтой до пурпурно-фиолетовой, коричневой и всех оттенков красного.
1. Светособирающие антенные комплексы – необходимы для улавливания света. Состоят из светособирающих пигментов, поглощающих энергию света, и передающие ее в реакционные центры. Функцию «антенны» выполняют пигменты: бактериохлорофиллыа, с, d, e, а также каротиноиды. У некоторых пурпурных бактерий, содержащих бактериохлорофилл в, он же выполняет функцию «антенны». Светособирающие пигменты организованы в виде комплексов с молекулами белков и локализованы или в ЦПМ и ее производных - тилакоидах (у пурпурных бактерий), или в хлоросомах (у зеленых бактерий), или в ЦПМ (у гелиобактерий).
2. Фотохимические реакционные центры (РЦ) – это трансмембранные белково-пигментные комплексы, локализованный в тилакоидах или ЦПМ. В РЦ происходит превращение электромагнитной формы энергии в химическую.
В состав РЦ входят первичный донор электронов, первичный акцептор электронов, вторичный акцептор электронов, вторичный донор электронов, а также каротиноиды и специфические белки.
Первичным донором электронов всегда является длинноволновая формабактериохлорофилла.
Первичный акцептор электроновразличен у разных групп фототрофных бактерий: может быть бхл с (у зеленых серных), бактериофеофитин а или в (у пурпурных серных и несерных).
Природа вторичного акцептора электроновтакже различна у разных групп: может быть убихинон, или менахинон (как у пурпурных бактерий) или железосероцентры (у зеленых серных бактерий). Их может быть один или более.
В роли вторичного донора электроноввыступают цитохромы типа с.
3.Третий компонент фотосинтетического аппарата – это фотосинтетические электронтранспортные системы. Обеспечивают перенос электронов, сопряженный с запасанием энергии в молекулах АТФ. Состоит из переносчиков электронов – цитохромов. Электронтранспортные цепи локализованы в мембранах (тилакоидах или ЦПМ).
В аноксигенном фотосинтезе участвует только одна световая реакция; она поддерживает циклический транспорт электронов. Электроны, покидающие цикл для восстановления NAD, не являются продуктом разложения воды. Фотосинтез зависит от наличия в среде восстановленных субстратов и не сопровождается выделением 02. Собственно фотореакция хотя и аналогична первой фотореакции у зеленых растений, однако у некоторых бактерий она приводит, вероятно, лишь к созданию протонного потенциала и тем самым к запасанию энергии (АТР), но не к восстановлению NAD. Таким образом, нециклический перенос электронов (от донора электронов к пиридиннуклеотиду) здесь отсутствует. По-видимому, NADH2 образуется в результате какой-то темновой реакции в ходе обратного транспорта электронов, протекающего с затратой энергии.
Следует, однако, заметить, что у фототрофных бактерий между отдельными группами существуют гораздо большие различия в составе пигментов и механизмах фотосинтеза, чем у зеленых растений. В последующем изложении мы вначале не будем касаться зеленых бактерий.
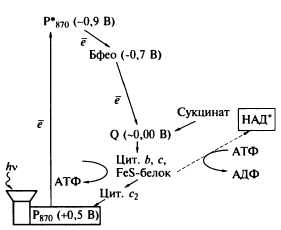
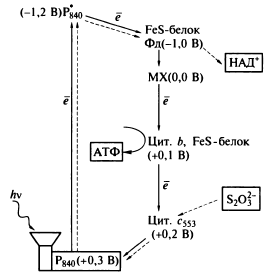
пурпурные - разнородная группа фотосинтезирующих протеобактерий, обитающих в солёных и пресных водах. Пурпурные бактерии относятся к классам альфа-, бета-, и гамма-протеобактерий.
Серные. Сюда относятся в основном представители Гаммапротеобактерий. Облигатные анаэробы. Прототрофы и фотолитоавтотрофы. Для родов Thiocapsa и Thiospirillum показана возможность хемолитоавтотрофного роста с молекулярным водородом и сульфидом в качестве доноров электронов. Подвижные формы обладают полярными жгутиками. Способность использовать органические вещества как доноры электронов ограничена. В основном они служат как источники углерода. могут сбраживать пируват до ацетата. Бактериохлорофиллы имеют максимумы поглощения 470 нм, 870 нм и 1070 нм.
Несерные. Сюда обычно относят представителей из порядков Rhodospirillales, Rhodobacterales и Rhodocyclales. Имеют ферменты защиты от O2. В микроаэрофильных и аэробных условиях способны расти в темноте. Имеют склонность к фотоорганогетеротрофному образу жизни, предпочитая в качестве доноров электронов и источника углерода органические вещества, однако могут быть и фотолитоавтотрофами. У подвижных форм есть жгутики, у многих присутствуют газовые вакуоли. Есть и неподвижные формы. Нуждаются в некоторых витаминах. Бактериохлорофиллы имеют максимумы поглощения 400 нм, 800 нм и 1030 нм.
место обитания. основном водные микроорганизмы. Помимо этого они встречаются в почве, но там их роль невелика. Развиваются обычно в бескислородных водах с сероводородом, куда проникает свет, в редких случаях обнаруживаются на бо́льших глубинах. Пурпурные несерные бактерии предпочитают богатые органикой воды и болотистые почвы, при этом редко образуют скопления, придающие воде окраску. Иногда развиваются в прибрежных морских водах. Пурпурные серные бактерии, наоборот, создают видимые скопления в прозрачных водоёмах на границе анаэробной зоны. Такие слои лучше всего формируются в меромиктических (с более высокой придонной солёностью) или голомиктических (с сезонной стратификацией) водоёмах и по берегам морей в лиманныхобластях. В прибрежных зонах морей могут образовывать красные приливы. Представители Ectothiorhodospira тяготеют к солёным и щелочным местообитаниям, морским эстуариям. В глобальном круговороте серы функционально тесно связаны с сульфатредукторами.
зеленые.
Серные. семейство облигатно анаэробных (более строгих, чем пурпурные бактерии, в присутствии O2 не растут)фотолитоавтотрофных грамотрицательных бактерий, использующих сероводород (H2S), водород (H2) и элементарную серу (S0) в качестве доноров электронов. По происхождению они принадлежат к надтипу Bacteroidetes-Chlorobi, однако неоднородны и потому их классифицируют как отдельный тип.
Зелёные серобактерии также обнаружены в Озере Матано, Индонезия, на глубине приблизительно 110—120 метров. Популяция, возможно, содержит видыChlorobium ferrooxidans.
Зелёные серобактерии редко образуют скопления. Они растут в илах в зоне хемоклина под слоем пурпурных бактерий. С глубиной зелёные виды замещаются коричневыми. Для них характерно образование консорциумов на основе синтрофии по сероводороду.
Галоархеи. одноклеточные микроорганизмы разной морфологии, часто плейоморфные, некоторые имеют газовые вакуоли. Клетки неподвижны или движутся с помощью лофотрихальных жгутиков. Пигментация галоархей часто бывает красно-желтой из-за большого количества каротиноидов, которые, возможно, используются микроорганизмами для защиты от излишней инсоляции. Характерной чертой этих микроорганизмов служит их абсолютная зависимость от высоких концентраций NaCl (до 20-30%).
Способность к фотосинтезу реализуется у галобактерий в неблагоприятных условиях при недостатке кислорода. В концентрированных растворах соли кислород очень плохо растворим, что приводит к созданию почти анаэробных условий. В этом случае галоархеи на свету при низком содержании кислорода образуют модифицированные клеточные мембраны, содержащие несколько видов пигментов – бактериородопсин, галородопсин, слоуродопсин, фобородопсин и археродопсин. Бактериородопсин отвечает за транслокацию протонов, галородопсин закачивает К+ в обмен на Na+, а слоу-, фобо- и археродопсины служат фоторецепторами на красный и синий свет, управляют активностью жгутиков для осуществления фототаксиса. Бактериородопсин(или ретиналь) синтезируется в присутствии небольшого количества О2 из С40-каротиноидов. Окисленный С20-фрагмент присоединяется через лизин к белковой молекуле и мигрирует в ЦПМ, образуя там агрегаты. Это соединение имеет темно-пурпурный цвет, поэтому агрегаты получили название «пурпурных мембран». При поглощении света бактериородопсин изменяет конформацию, что сопровождается выбросом протонов из клетки и наведением трансмембранного потенциала. Энергия ΔμН+ с помощью фермента АТФазы преобразуется в макроэргические связи. Таким образом, галобактерии используют световую энергию, чтобы в неблагоприятных условиях синтезировать достаточное количество АТФ и пережить неблагоприятные условия.
Поможем написать любую работу на аналогичную тему
Реферат
Бактериальный фотосинтез. Оксигенные и аноксигенные фототрофные бактерии. Пигменты аноксигенных фототрофных бактерий.
От 250 руб
Контрольная работа
Бактериальный фотосинтез. Оксигенные и аноксигенные фототрофные бактерии. Пигменты аноксигенных фототрофных бактерий.
От 250 руб
Курсовая работа
Бактериальный фотосинтез. Оксигенные и аноксигенные фототрофные бактерии. Пигменты аноксигенных фототрофных бактерий.
От 700 руб