Энергетический метаболизм (катаболизм) – это совокупность реакций окисления различных восстановленных органических и неорганических соединений, сопровождающихся выделением энергии, аккумулируемой клеткой в форме фосфатных связей.
Хемотрофные микроорганизмы используют для синтеза молекул АТФ энергию, освобождаемую в результате химических реакций, фототрофные – световую энергию в процессе протекания фотосинтеза.
Синтез молекул АТФ из АДФ и фосфатов может происходить двумя способами:
• фосфорилированием в дыхательной или фотосинтетической электронтранспортной цепи. (мембранным фосфорилированием) Синтез АТФ в данном случае происходит при участии фермента АТФ-синтазы:
• фосфорилированием на уровне субстрата. При этом фосфатная группа переносится на АДФ от вещества (субстрата), более богатого энергией, чем АТФ.
У хемотрофных бактерий генерация энергии в молекулах АТФ сводится к двум типам биохимических реакций: окисления и восстановления.
При биологическом окислении чаще всего происходит одновременный перенос двух электронов; при этом от субстрата отщепляются также два протона (Н+) (дегидрирование).
Окислительно-восстановительные реакции энергетического метаболизма у хемотрофных:
• аэробное дыхание, или аэробное окисление; • анаэробное дыхание; • брожение.
аэробное дыхание, при котором донором водорода или электронов являются органические (реже неорганические) вещества, а конечным акцептором – молекулярный кислород. Основное количество энергии при аэробном дыхании образуется в электронтранспортной цепи, т. е. в результате мембранного фосфорилирования.
Анаэробное дыхание – цепь анаэробных окислительно-восстановительных реакций, которые сводятся к окислению органического или неорганического субстрата с использованием в качестве конечного акцептора электронов не молекулярного кислорода, а других неорганических веществ (нитрата – , нитрита – , сульфата – , сульфита), а также органических веществ (фумарата и др.). Молекулы АТФ в процессе анаэробного дыхания образуются в основном в электронтранспортной цепи, т. е. в результате реакций мембранного фосфорилирования, но в меньшем количестве.
Брожение – совокупность анаэробных окислительно-восстановительных реакций, при которых органические соединения служат как донорами, так и акцепторами электронов. АТФ при брожении синтезируется в результате реакций субстратного фосфорилирования.
У бактерий возможны три пути катаболизма глюкозы:
1) гликолиз, или фруктозодифосфатный путь, или путь Эмбдена – Мейергофа – Парнаса (по имени исследователей, внесших большой вклад в изучение этого процесса);
2) окислительный пентозофосфатный путь, или гексозомонофосфатный путь, или путь Варбурга – Диккенса – Хореккера;
3) 2-кето-3-дезокси-6-фосфоглюконатный путь (КДФГ-путь), или путь Энтнера – Дудорова.
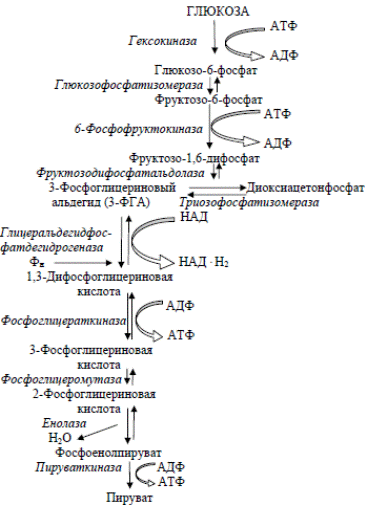
Глюкоза сначала фосфорилируется при участии фермента гексокиназы и АТФ как донора фосфата. Образуется глюкозо-6-фосфат.
Гликолиз. С6Н12О6 + 2АДФ + 2Фн + 2НАД = 2С3Н4О3 + 2АТФ + 2НАД · Н2
Пентозофосфатный путь (семейства Enterobacteriaceae,гетероферментативных молочнокислых бактерий и некоторых маслянокислых бактерий). В этом цикле глюкозо-6-фосфат, образующийся путем
активирования глюкозы молекулой АТФ, превращается через ряд промежуточных реакций в 6-фосфоглюконовую кислоту, которая подвергается окислению и декарбоксилированию с образованием рибулозо-5- фосфата, СО2 и НАДФН2 . Рибулозо-5-фосфат включается в сложный цикл, приводящий к образованию из трех его молекул двух молекул глюкозо-6-фосфата и одной молекулы 3-фосфоглицеринового альдегида. Глюкозо-6-фосфат может снова включаться в цикл, а 3-ФГА может быть превращен в пировиноградную кислоту.
 Одна молекула АТФ. Он обеспечивает клетки бактерий пентозами (рибулозо-5-фосфатом). Образуются две молекулы НАДФН2.
Одна молекула АТФ. Он обеспечивает клетки бактерий пентозами (рибулозо-5-фосфатом). Образуются две молекулы НАДФН2.
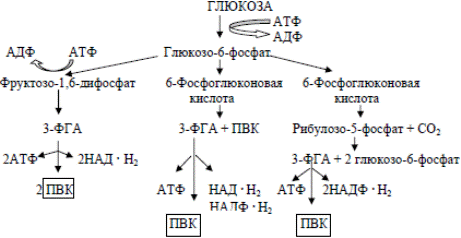 Путь Энтнера – Дудорова характерен в основном для псевдомонад и уксуснокислых бактерий. 6-фосфоглюконовая кислота превращается в ПВК и 3-ФГА. Последний может превращаться в пировиноградную кислоту. Из одной молекулы глюкозы при функционировании этого пути синтезируется одна молекула АТФ, по одной молекуле НАДФН2 и НАДН2. Кратчайшим механизмом расщепления углеводов ПВК. Аэробное дыхание
Путь Энтнера – Дудорова характерен в основном для псевдомонад и уксуснокислых бактерий. 6-фосфоглюконовая кислота превращается в ПВК и 3-ФГА. Последний может превращаться в пировиноградную кислоту. Из одной молекулы глюкозы при функционировании этого пути синтезируется одна молекула АТФ, по одной молекуле НАДФН2 и НАДН2. Кратчайшим механизмом расщепления углеводов ПВК. Аэробное дыхание
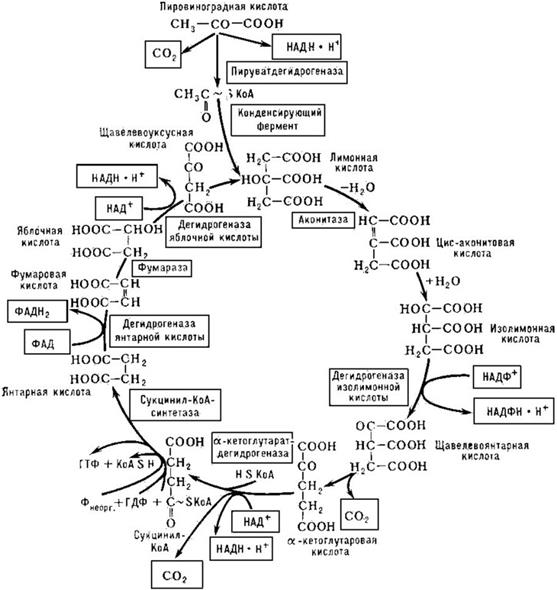
Пвк окисляется с участием коэнзима А до ацетил-КоА. В данном процессе работают ферменты пируватдегидрогеназы: CH3–CO–COOH + KoA–SH + HAД+ = CH3–CO~KoA + НАД · Н2 + CO2 Ацетил-КоА является исходным субстратом цикла трикарбоновых кислот (ЦТК), или цикла Кребса.
В конечном итоге окисление ацетил-КоА в ЦТК приводит к образованию двух молекул СО2, одной молекулы АТФ и восьми атомов водорода, из которых шесть атомов связаны в молекулах пиридиннуклеотидов и два атома – в молекулах флавопротеинов.
У некоторых бактерий цикл трикарбоновых кислот «разорван». Отстутствует этап превращения а-кетоглутаровой кислоты в янтарную. В таком виде ЦТК не может функционировать в системе энер-
годающих реакций клетки. Основная функция «разорванного» ЦТК – биосинтетическая.
Образовавшиеся на разных этапах окисления органических веществ НАДН2 и ФАДН2, поступают в дыхательную цепь, которая у бактерий находится в цитоплазматической мембране. Они вновь окисляются до НАД и ФАД, а отщепившийся от них водород передается не менее чем через пять переносчиков на заключительный участок цепи, где соед-ся с молекулярным кислородом, образуя воду.
Транспорт водорода сопровождается протеканием ряда окислительно-восстановительных реакций. Достаточно энергии для образования АТФ, процесс окислительного фосфорилирования. Флавопротеины и хиноны осуществляют перенос атомов водорода, а FeS-белки и цитохромы – электронов. Энергетическю выход при окислении одной мол-лы глюк. при аэробн. дыхании у дрожжей: • в процессе гликолиза - по 2 АТФ, 2 НАДН2 и 2 пирувата; • при окислительном декарбоксилировании 2 пирувата образуются 2 ацетил-КоА и 2 НАДН2; • окисление двух молекул ацетил-КоА в цикле Кребса приводит к образованию 6 НАДН2, 2 ФАДН2 и 2 АТФ. При окислении одной молекулы НАДН2 = 3 АТФ, ФАДН2 = 2 АТФ. При окислении 10 молекул НАДН2 = 30 молекул АТФ, а двух молекул ФАДН2 = 4 молекулы АТФ. Суммарный энергетический выход аэробного дыхания у эукариотических микроорганизмов, когда катаболизм глюкозы осуществляется гликолитическим путем, составляет 38 молекул АТФ: С6Н12О6 + 6О2 + 38АДФ + 38Н3РО4 6СО2 + 38АТФ + 44Н2О
 E. coli, когда ката-
E. coli, когда ката-
болизм глюк.происх. гликолитич. путем, образуется 26 АТФ:
• 2 АТФ синтезир. в гликолизе;
• 2 АТФ синтезир. в двух оборотах цикла Кребса;
• 10 молекул НАДН2 приводят к синтезу 20 молекул АТФ;
• две молекулы ФАДН2 = 2 АТФ.
У других прокариот, таких как Corynebacterium diphtheriae, в дыха-
тельной цепи имеется только один пункт «выброса» протонов. У Mycobacterium
phlei – три, как в дыхательной цепи митохондрий дрожжей.
Бактерии, способные к анаэробному дыханию, имеют укороченные дыхательные цепи. В дыхательных цепях анаэробов цитохромоксидаза заменена соответствующими редуктазами. У строгих анаэробов не функционирует цикл Кребса или же он разорван.Основное количество молекул АТФ при анаэробном дыхании синтезируется в процессе мембранного фосфорилирования. Выход АТФ при анаэр.дых. меньше,чем при аэр-ом,но больше, чем при брожении. Конечным акцепторами электронов при нитратном дыхании являются нитраты или нитриты. Результатом нитратного дыхания является восстановление до газообразных продуктов. Суммарную реакцию нитратного дыхания, где окисляемым субстратом является глюкоза, а конечным акцептором электронов – нитраты: C6H12O6 + 4NO3=6CO2+6H2O + 2N2 + х (кДж) У денитрифицирующих бактерий функционирует полная и укороченная электр.цепь. В качестве конечного акцептора электронов выступает сульфат (SO4), в результате чего происходит его восстановление до Н2S. Получение энергии в результате сульфатного дыхания состоит из трех этапов • отрыва электронов от энергетического субстрата; • переноса их по дыхательной цепи• присоединения их к веществам, функционирующим в качестве конечных акцепторов электронов. Цикла Кребса «разорван» и функционирует только в условиях конструктивного метаболизма. Карбонатное дыхание – процесс окисления молекулярного водорода, при котором конечным акцептором электронов является СО2. Метаногенные бактерии в основном получают энергию за счет окисления молекулярного водорода в процессах, сопряженных с восстановлением СО2: 4Н2 + СО2 = СН4 + 2Н2О Многие метаногенные бактерии могут испол-вать для получен. энергии формиат, метанол, ацетат, а также метилированные амины: 4НСООН = СН4 + 3СО2 + 2Н2О, 4СН3ОН = 3СН4 + СО2 + 2Н2О, 4СО + 2Н2О = СН4 + 3СО2, СН3СООН = СН4 + СО2, 4СН3NH3 + 2H2O = 3CH4 + CO2 + 4NH4.
Восстановление фумарата в бактериальных клетках часто сопровождается образованием сукцината (Bacteroides, Fibrobacter, Wolinella). Кроме сукциногенных бактерий, к фумаратному дыханию способны многие другие хемоорганотрофы (энтеробактерии (роды Escherichia, Proteus, Salmonella).
По своей биологической сути брожение – это способ получения энергии, при котором АТФ образуется в результате анаэр. окисления органических субстратов в реакциях субстратного фосфорилирования.
Сиртовое. Катаболизм глюкозы проходит по гликолитич. пути до стадии синтеза пировиноградной кислоты. Далее осуществляется ее декарбоксилирование пируватдекарбоксилазой при участии тиаминпирофосфата, в результате чего образуются ацетальдегид и СО2. Ацетальдегид выступает конечным акцептором водорода. При помощи алкогольдегидрогеназы он восстанавливается до этанола.
C6H12O6 + 2Фн + 2АДФ = 2CH3CH2OH + 2CO2 + 2АТФ + 2H2O
Виды дрожжей (Saccharomyces cerevisiae, S. uvarum, Schizosaccharomyces pombe и др.) и бактерий (Erwinia amylovora, Sarcina ventriculi, Zymomonas mobilis). Кроме того, этанол образуют такие мезофильные бактерии, как Leuconostoc mesenteroides, Clostridium sporogenes, Spirochaeta aurantia, а также термофильные бактерии Thermoanaerobacter ethanolicus, Clostridium thermohydrosulfuricum.
Маслянокислое проходит в строго анаэробных условиях и осуществляют его облигатно-анаэробные бактерии рода Clostridium. Сбраживание углеводов по гликолитическому пути. Основной продукт брожения – масляная кислота синтезируется в результате конденсации двух молекул ацетил-КоА. Кроме того, одна из молекул ацетил-КоА, присоединяя неорганический фосфат, может подвергаться фосфорилированию, превращаясь в ацетилфосфат и далее в ацетат, что сопровождается синтезом АТФ при субстратном фосфорилировании. Это третья молекула АТФ, которая синтезируется при маслянокислом брожении (две другие молекулы АТФ образуются при расщеплении глюкозы по гликолитическому пути). Расчеты показали, что в целом на одну молекулу сбраживаемой глюкозы в маслянокислом брожении образуется 3,3 молекулы АТФ.
При гомоферментативном молочнокислом брожении синтезируется практически одна молочная кислота (90 % всех продуктов брожения). Катаболизм глюкозы в этом случае происходит по гликолитическому пути. Образ-яся при ПВК не подвергается декарбоксилированию, а под действием лактатдегидрогеназы восстанавливается до молочной к=ты. Конечным акцептором водорода выступает ПВК.
С6Н12О6 + 2АДФ + 2Фн = 2СН3 – СНОН – СООН + 2АТФ. Возбудителями гомоферментативного молочнокислого брожения являются, например, бактерии Streptococcus cremoris, S. lactis, Lactobacillus
bulgaricus, L. lactis и др.
Гетероферментативное молочнокислое брожение приводит к образованию разнообразных продуктов: молочной и уксусной кислот, этилового спирта, углекислого газа и глицерина. При этом типе брожения расщепление углеводов происходит по пентозофосфатному пути. Конечными акцепторами водорода являются ПВК и ацетальдегид. Возбудителями гетероферментативного молочнокислого броже-
ния являются бактерии видов Leuconostoc mesenteroides, Bifidobacterium bifidum, Lactobacillus brevis.
Основным продуктом, образующимся при пропионовокислом брожении, является пропионовая кислота. Кроме нее, синтезируются уксусная кислота и СО2. 1,5Глюкоза = 2Пропионат + СО2 + Ацетат
Акрилатный путь (Clostridium propionicum, Bacteroides ruminicola, Megasphaera elsdenii).
Выход АТФ составляет одну молекулу на три молекулы потребленного в этом случае лактата.
Сукцинат-пропионатный функционирует у большинства микроорганизмов, образующих пропионат.
На этапе превращения фумарата в сукцинат происходит субстратное фосфорилирование, в результате чего образуется молекула АТФ.
смешанного типа 1. Брожение, характерное для бактерий родов Escherichia, Salmonella, Shigella, Citrobacter, Yersinia, при котором образуются главным образом кислоты (молочная, уксусная, янтарная, муравьиная). Выход АТФ в этом случае составляет 2–2,5 молекулы АТФ на одну молекулу глюкозы. (АТФ синтезируется в реакции, катализируемой ацетаткиназой)
Поможем написать любую работу на аналогичную тему