Классификация – распределение множества организмов по группам (таксонам).
Номенклатура – присвоение названий отдельным группам и видам микроорганизмов.
Основной таксономической категорией является вид. Виды объединяются в роды, роды – в семейства, семейства – в порядки, далее следу-ют классы, отделы, царства. В микробиологии существуют также более мелкие таксономические единицы, чем вид: подвид (subspeciens), разновидность. Подвиды могут различаться по физиологическим (biovar), морфологическим (morphovar) или по антигенным (serovar) свойствам. Большое значение в микробиологии имеют такие понятия, как клон – чистая культура, полученная из одной клетки, и штамм – культуры бактерий одного вида, выделенные из различных источников либо из одного источника в разное время или полученные в ходе генетических манипу-
ляций.
В настоящее время в микробиологии приняты два различных подхода к систематике, обусловливающих существование двух систем классификации: филогенетической и фенотипической. В основу филогенетической классификации положена идея создания системы прокариот, объективно отражающей родственные отношения между разными группами бактерий и историю их эволюционного развития. Фенотипическая классификация преследует, в первую очередь, практические цели, заключающиеся в том, чтобы быстрее установить принадлежность микроорганизма к определенному таксону. Определителе бактерий Берджи В конце 50-х годов ХХ в. возникла нумерическая (численная) таксономия французского ботаника М. Адансона (1757).
При идентификации бактерий приоритетным является использование генетических (молекулярно-биологических), фенотипических и серологических подходов и критериев систематики.
Наиболее объективными и дающими представление о филогенетических связях являются генетические (молекулярно-биологические) критерии. К ним относятся определение относительного содержания ГЦ-пар в ДНК, нуклеотидных последовательностей в молекулах ДНК или РНК, гибридизация нуклеиновых кислот, применение генетических зондов, рестрикционный анализ ДНК, методы генетического анализа (изучение переноса генов, генетических скрещиваний, картирование хромосом).
Близкородственные микроорганизмы имеют идентичное или сходное содержание ГЦ-пар в ДНК, а далеко отстоящие в генетическом отношении сильно отличаются по относительному содержанию этих азотистых оснований.
Нуклеотидный состав ДНК бактерий можно определить химическими и физическими методами.
К химическим относится метод хроматографии на бумаге: выделение ДНК, ее гидролиз до азотистых оснований, разделение их с помощью хроматографии на бумаге, элюирование оснований с бумаги и
последующая ультрафиолетовая спектрофотометрия.
К физическим относятся метод определения содержания азотистых оснований по температуре плавления ДНК и метод ультрацентрифугирования ДНК в градиенте плотности хлорида цезия.
Температура плавления – это темп., при которой происходит денатурация ДНК в результате разрыва водор. связей между азотист.осн. Чем выше содержание ГЦ-пар в ДНК, тем выше темп. ее плавления.
 Разделение цепей сопровождается увеличением оптической плотности при 260 нм, т. е. максимуме поглощения ДНК в УФ-свете, что легко измерить спектрофотометрически. При постепенном нагревании ДНК поглощение увелич. по мере разрыва водородных связей и достигает плато при температуре, когда ДНК становится полностью одноцепочечной. Средняя точка на кривой – температура плавления (Тпл.) – служит мерой содержания ГЦ-оснований, а нуклеотидный состав ДНК: (Г + Ц) % = (Тпл. – 69,3°) * 2,439.
Разделение цепей сопровождается увеличением оптической плотности при 260 нм, т. е. максимуме поглощения ДНК в УФ-свете, что легко измерить спектрофотометрически. При постепенном нагревании ДНК поглощение увелич. по мере разрыва водородных связей и достигает плато при температуре, когда ДНК становится полностью одноцепочечной. Средняя точка на кривой – температура плавления (Тпл.) – служит мерой содержания ГЦ-оснований, а нуклеотидный состав ДНК: (Г + Ц) % = (Тпл. – 69,3°) * 2,439.
Метод ультрацентрифуг. в град. плотности хлорида цезия основан на том, что имеется линейная зависимость между плотностью ДНК и содержанием в ней ГЦ-пар.
Существуют и другие методы (при помощи бромирования оснований, депуринизации, спектраль-
ного анализа, электрофореза). Более тонким методом оценки генетического сходства организмов является метод молекулярной гибридизации нуклеиновых кислот, с помощью которого определяют число и степень сходства гомологичных участков в геномах сравниваемых видов. В основе метода лежит способность денатурированных (одноцепочечных) ДНК в подходящих условиях реассоциировать, соединяться с образованием двухцепочечных молекул ДНК.
Простейший повсеместно используемый способ изучения реассоциации нуклеиновых кислот – метод с применением колонки, содержащей гидроксилапатит. Гидроксилапатит представляет собой гель фосфата кальция, который при определенных условиях специфически адсорбирует только двойные, но не одиночные цепи нуклеиновых кислот.
Образование генетических рекомбинантов служит значительно более точным показателем уровня генетической гомологии, чем гибридизация in vitro, поскольку включение каждого отдельного фрагмента молекулы ДНК донора зависит от степени его гомологии с ДНК реципиента именно в том небольшом специфическом участке хромосомы, в котором должна произойти рекомбинация.
Ферменты рестриктазы способны распознавать специфические нуклеотидные последовательности и в строго определенных участках (сайтах рестрикции) «разрезать» молекулы ДНК на фрагменты (рестрикты). Расположение фрагментов ДНК, продуктов расщепления, разделенных с помощью электрофореза в агарозном геле, дает существенную информацию о типе и количестве специфических нуклеотидных послед-стей в хромосомах и позволяет судить об их сходстве или различии орг-ов.
Метод молекулярных зондов (ДНК-зондов) основан на реакции гибридизации между фрагментом нуклеотидной последовательности (зондом), несущим наиболее специфический и консервативный для данного вида бактерий ген, с полимерной ДНК изучаемого микроорганизма.
В основе ПЦР лежит многократное реплицирование специфического участка нуклеотидной послед-сти, катализируемое ДНК-зависимой ДНК-полимеразой, и использование праймера – фрагмента ДНК, несущего наиболее специфичную для данного микроорг. нуклеотидную послед-сть гена (участка гена).
Определение нуклеотидных последовательностей (секвенирование) дает возможность проводить сопоставительный анализ последовательностей в различных молекулах ДНК и РНК. Чаще всего анализируются 16S-рРНК.
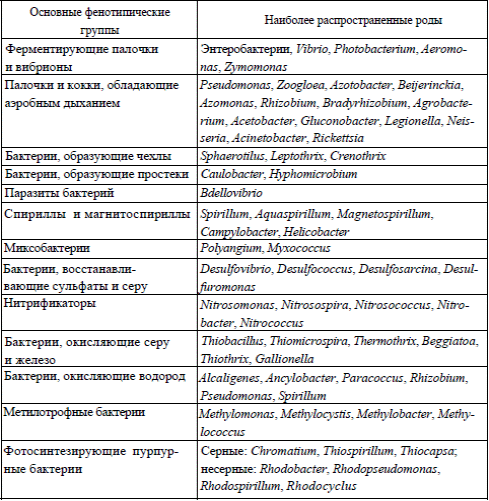
В классификации бактерий используют набор фенотипических признаков: морфологических, культуральных, физиологических и биохимических.
Серологические (от лат. serum – сыворотка) критерии систематики основаны на специфических реакциях взаимодействия антигенов (компоненты клеточных стенок, жгутиков, капсул, ДНК и токсинов) идентифицируемых микроорганизмов с антителами, содержащимися в сыворотках.
Для постановки серологических реакций необходима сыворотка, которую получают из крови лабораторного животного, иммунизированного коллекционным (известной видовой и штаммовой принадлежности) микроорганизмом. Она содержит антитела, специфичные к данному штамму.
Приемлемой филогенетической системой классификации прокариот является система, основанная на сопоставлении последовательности нуклеотидов в 16S-рРНК. Все прокариоты разделены на 26 филогенетических «ветвей»; 23 «ветви» представлены эубактериями, а три – архебактериями.
Из 23 групп - 2 филогенетические группы представлены грампол. бактериями, остальные – грамотр.
Грамотрицательные бактерии состоят из крупной группы Протеобактерий (Proteobacteria) и 20 групп
 Протеобактерии – очень гетерогенная группа. Клетки большинства видов. Разделена на пять подгрупп: альфа, бета, гамма, дельта и эпсилон.
Протеобактерии – очень гетерогенная группа. Клетки большинства видов. Разделена на пять подгрупп: альфа, бета, гамма, дельта и эпсилон.
К грамотрицательным относятся следующие группы эубактерий: водородные термофилы, зеленые нитчатые бакт., зеленые серные бакт., цианобакт., спирохеты, цитофаги, хламидии.
Филогенетические группы грамположительных бактерий – Actinobacteria и Firmicutes. Группа Actinobacteria («актиномицетная ветвь») представлена родами бактерий, имеющими в ДНК высокое содержание ГЦ-пар: Geodermatophilus,Frankia, Strepto myces, Arthrobacter,Micrococcus, Actinomyces,Bifidobacterium, Propionibacterium, Actinoplanes, Nocardia, Rhodococcus, Corynebacterium, Mycobacterium. Группа Firmicutes («клостридиальная ветвь») состоит из: Clostridium, Lactococcus, Pediococcus, Streptococcus, Enterococcus, Leuconostoc, Listeria, Caryophanon, Staphylococcus, Sarcina, Sporosаrcina, Bacillus, Desulfotomaculum, Heliobacterium, Mycoplasma, Ureaplasma.
В составе архебактерий выделяют три филогенетические группы: Crenarchaeota, Euryarchaeota и Korarchaeota. Группа Crenarchaeota состоит из экстремально термофильных бактерий, большинство которых осуществляют метаболизм серы, некоторые восстанавливают ионы железа и молибдена. В группу Euryarchaeota входят облигатно анаэробные метаногенные архебактерии, а также экстремальные термофилы и галофилы. Группа Korarchaeota образована архебактериями,обитающими в горячих серных источниках.
Наиболее признанной и используемой фенотипической классификацией бактерий является классификация, представленная в девятом издании Определителя бактерий Берджи:
1) Gracilicutes – грамотрицательные эубактерии, имеющие клеточные стенки; 2) Firmicutes – грамположительные эубактерии, имеющие клеточные стенки; 3) Tenericutes – эубактерии, лишенные клеточных стенок; 4) Mendosicutes – архебактерии, клеточные стенки которых отличаются.
В отдел Gracilicutes входят бактерии различной морфологии с грамотрицательной клеточной стенкой. Размножение происходит в основном бинарным делением, некоторые бактерии размножаются почкованием. Эндоспор не образуют. Большинство подвижны: встречаются все типы передвижения бактерий – с помощью жгутиков, скольжением, изгибанием. Отдел включает аэробные, анаэробные и факультативные анаэробные бактерии; фототрофные и хемотрофные бактерии. Отдел подразделяют на три класса: Scotobacteria, Oxyphotobacteria, Anoxyphotobacteria. В класс Scotobacteria входят грамотрицательные бактерии, не использующие световую энергию для целей метаболизма, а получающие ее только в результате окислительно-восстановительных реакций. В класс Anoxyphotobacteria входят пурпурные бактерии, зеленые бактерии и гелиобактерии, осуществляющие аноксигенный фотосинтез. Класс Oxyphotobacteria представлен цианобактериями и прохлорофитами, осуществляющими оксигенный фотосинтез.
В отдел Firmicutes включены бактерии с грамположительной клеточной стенкой. Клетки могут иметь разную форму: палочки, кокки, нитевидные, ветвящиеся. Некоторые представители образуют эндоспоры. Большинство из них неподвижны; подвижные формы имеют перитрихиальное жгутикование. В состав отдела входят аэробные, анаэробные и факультативно анаэробные бактерии. Отдел состоит из двух классов: Firmibacteria, Thallobacteria. Класс Firmibacteria включает большое ко-
личество «неветвящихся» бактерий. Класс Thallobacteria включает бактерии, к-ки которых «ветвяться».
Отдел Tenericutes представлен бактериями, не имеющими клеточной стенки. Форма клеток непосто-
янна. Размножение бактерий, входящих в этот отдел, происходит бинарным делением, почкованием. Окрашивание по Граму отрицательное. Хар-но образование мелких, врастающих в агар колоний. Могут быть сапрофитными, паразитами или патогенами. Отдел состоит из класса Mollicutes (микоплазмы).
Отдел Mendosicutes образован бактериями с ригидной клеточной стенкой, но не содержащей пептидогликана муреина. Большинство представителей – строгие анаэробы, многие из которых имеют жгутики. Виды характеризуются экологическим и метаболическим разнообразием, способностью жить в экстремальных условиях. Отдел состоит из одного класса – Archaebacteria.
К отделу Gracilicutes принадлежат следующие группы. 1. Спирохеты. 2. Аэробные, подвижные, спиралевидные грамотрицательные бактерии. 3. Неподвижные или редко подвижные грамотрицательные изогнутые бактерии. 4. Грамотрицательные аэробные (или микроаэрофильные) палочки и кокки. 5. Факультативно аэробные грамотрицательные палочки. 6. Грамотрицательные анаэробные прямые, изогнутые или спиралевидные палочки. 7. Бактерии, осуществляющие диссимиляционное восстановление серы или сульфата. 8. Анаэробные грамотрицательные кокки.
9. Риккетсии и хламидии. 10. Аноксигенные фототрофные бактерии. 11. Оксигенные фототрофные бактерии.12. Аэробные хемолитотрофные бактерии и близкие организмы. 13. Почкующиеся и (или) образующие выросты бактерии. 14. Бактерии, имеющие чехлы. 15. Нефотосинтезирующие скользящие бактерии, не образующие плодовых тел. 16. Скользящие бактерии, образующие плодовые тела.
В отдел Firmicutes входят: 17. Грамположительные кокки. 18. Грамположительные палочки и кокки, образующие эндоспоры. 19. Грамположительные палочки правильной формы, не образующие спор.
20. Грамположительные палочки неправильной формы, не образующие спор. 21. Микобактерии.
22–29. Актиномицеты.
К отделу Tenericutes принадлежит: Группа 30. Микоплазмы.
Отдел Mendosicutes включает 31. Метаногены. 32. Сульфатредуцирующие архебактерии.
33. Экстремально галофильные архебактерии (галобактерии). 34. Архебактерии, лишенные клеточной стенки. 35. Экстремально термофильные и гипертермофильные архебактерии, метаболизирующие серу.
Поможем написать любую работу на аналогичную тему