Впервые стали известны в 1977 г. благодаря работам американских ученых К. Везе и Г. Фокса по изучению молекулярно-биохимических свойств биополимеров клеток разных видов бактерий.
1. Их клеточная стенка не имеет пептидогликана муреина, вместо ко-торого в состав клеточной стенки входят кислые полисахариды, белки или псевдомуреин, не содержащий в отличие от муреина мурамовой кислоты, а в пептидных мостиках – D-аминокислот. Вместо ацетилмурамовой кислоты в состав муреина входит ацетилталозаминуроновая кислота, что определило устойчивость архебактерий к антибиотикам, нарушающим синтез клеточных стенок у эубактерий, – пенициллину, ампициллину.
2. Мембраны архебактерий не содержат в составе липидов сложных эфиров глицерина и жирных кислот, а представлены особыми бифитанильными глицериновыми эфирами, образованными путем конденсации глицерина с терпеноидными спиртами.
3. В тРНК архебактерий изменена общая для всех других организмов петля тимин–псевдоуридин–цитидин, в которой вместо тимина присутствуют другие основания.
4. Наличие в генах, кодирующих тРНК, интронов, которые имеются только в эукариотических геномах, но отсутствуют у большинства эубактерий.
5. Наличие в геноме архебактерий многократно повторяющихся последовательностей, что характерно для хромосомной ДНК эукариот. В области нуклеоида у архебактерий содержатся белки гистоны.
6. Архебактерии имеют более сложную структуру аппаратов трансляции и транскрипции. ДНК-зависимая РНК-полимераза, осуществляющая процесс транскрипции у архебактерий, состоит из 9–12 субъединиц, у эубактерий – из 4–8 субъединиц. РНК-полимераза архебактерий, подобно таковым у эукариот, не ингибируются рифампицином, их активность стимулируется силибином.
7. Рибосомы архебактерий содержат относительно больше белков, чем рибосомы эубактерий, причем они представлены более кислыми формами по сравнению с белками рибосом эубактерий. Кроме того, процесс биосинтеза белков у архебактерий не ингибируется такими антибиотиками, как хлорамфеникол и стрептомицин.
8. Особенностью конструктивного метаболизма архебактерий является отсутствие фиксации СО2 в цикле Кальвина. Основным путем автотрофной его фиксации является восстановительный путь карбоновых кислот в различных его модификациях, присущий и некоторым эубактериям.
9. Архебактерии неспособны использовать сложные высокомолекулярные соединения. Среди них не обнаружено активных продуцентов гидролитических ферментов, что, возможно, является одной из причин отсутствия патогенных и паразитических форм.
10. Некоторые архебактерии, в частности метаногенные, синтезируют уникальный набор коферментов, не встречающийся у других организмов
11. Архебактерии занимают необычные, часто экстремальные по условиям окружающей среды высокоспециализированные экологические ниши.
12. Морфология клеток архебактерий беднее, чем эубактерий. Преобладают сферические и цилиндрические клетки, а также необычные плоские клетки, имеющие вид пластинок и коробочек разнообразной геометрической формы, сходные с кусочками битого стекла.
Метаногенные бактерии – самая многочисленная группа, являющаяся облигатными анаэробами. Большинство неподвижны, подвижные имеют полярные жгутики. Источниками энергии служат процессы окисления молекулярного водорода, оксида углерода, метанола, муравьиной и уксусной кислот, акцептором электронов является углекислый газ. В природных средах метаногенные бактерии развиваются в ассоциации с другими микроорганизмами, выполняя функцию конечного звена в трофической цепи – превращают продукты брожения этих микроорганизмов в метан. Основные места обитания: торфяные болота, ил на дне водоемов, очистные сооружения сточных вод, пищеварительный тракт животных. Представители метаногенных бактерий входят в роды Methanobacterium, Methanosarcina, Methanospirillum.
Анаэробные серовосстанавливающие бактерии представлены одним родом Archaeoglobus, который состоит из двух видов: A. fulgidus (типовой вид) и A. profundus. Это облигатные анаэробы и экстремальные термофилы. Бактерии рода Archaeoglobus – грамотрицательные кокковидные клетки неправильной формы, часто треугольные, одиночные или в парах, со жгутиками (монополярные политрихи) или без них. При освещении светом с длиной волны 420 нм обнаруживают голубовато-зеленую флуоресценцию. На агаризованной среде формируют зеленовато-черные гладкие колонии диаметром 1–2 мм. Способны к хемолитотрофному или хемоорганотрофному росту. Основная форма энергетического метаболизма – анаэробное дыхание (диссимиляционная сульфатредукция). Донорами электронов являются формиат, лактат, глюкоза, крахмал, белки и молекулярный водород; конечными акцепторами электронов – сульфат, сульфит и тиосульфат (но не молекулярная сера). Особенностью бактерий рода Archaeoglobus является способность в небольшом количестве образовывать метан.
Основное местообитание серовосстанавливающих архебактерий – мелководные и глубоководные морские гидротермальные источники, в которых они вызывают восстановление соединений серы.
Экстремальные термофилы, метаболизирующие молекулярную серу – грамотрицательные бактерии разной морфологии: кокки, палочки, диски, нити или клетки неправильной дольчатой формы. Клеточные стенки у этих бактерий состоят из гликопротеиновых или белковых субъединиц. Цитоплазматические мембраны многослойные, содержат липиды, на основе тетраэфиров глицерина.
Экстремальные термофилы, метаболизирующие молекулярную серу, подразделяются на три порядка, четыре семейства и включают девять родов. Все представители объединены в одну группу благодаря тому, что их энергетический метаболизм связан с метаболизмом молекулярной серы. Облигатно аэробные бактерии (например, бактерии рода Sulfolobus); строгие анаэробы (например, бактерии ро-
дов Thermococcales и Thermoproteales); факультативные анаэробы (например, бактерии рода Acidianus).
Являются аборигенами высокотермальных кислых источников и грунтов в зонах вулканического происхождения. Кроме того, бактерии рода Pyrodictium выделяют из подводных морских горяч.источн.
Бактерии вида Sulfolobus brierley, способные выщелачивать металлы
Термоацидофильные микоплазмы представлены единственным видом Thermoplasma acidophilum. В отличие от других архебактерий, эти бактерии не имеют клеточной стенки. Клетки окружены трехслойной мембраной, толщиной около 7 нм, могут быть подвижными и обладать жгутиками.
Гетеротрофы со сложными пищевыми потребностями. Хемоорганотрофы, факультативные анаэробы. Энергию получают как за счет аэробного дыхания, так и за счет брожения. Облигатные термофилы и облигатные ацидофилы. Естественным местообитанием их служат саморазогревающиеся отходы каменного угля и кислые термальные источники.
В группу экстремально галофильных бактерий входят бактерии с разной морфологией клеток.
Галобактерии распространены там, где есть: высокое содержание NaCl и других необходимых ионов,
. У представителей рода Halobacterium клеточная стенка построена из регулярно расположенных гексагональных субъединиц, состоящих в основном из гликопротеинов. Клеточная стенка галобактерий рода Halococcus имеет гетерополисахаридную природу.
Цитоплазматическая мембрана галобактерий содержит липиды, в молекулах которых глицерин связан не с остатками жирных кислот, а с С20- терпеноидным спиртом – фитанолом. Включают много каротиноидных пигментов (основной – бактериоруберин)против избыточной радиации,.
При недостатке в среде молекулярного кислорода в цитоплазматической мембране галобактерий индуцируется синтез хромопротеина – бактериородопсина, белка, соединенного ковалентной связью с каротиноидом ретиналем.Хромопротеин откладывается в виде отдельных пурпурных областей
(бляшек) красного цвета на цитоплазматической мембране.
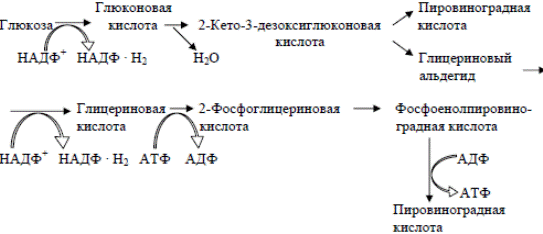
Экстремальные галофилы имеют сложные пищевые потребности. Метаболизм глюкозы осуществляется по модифицированному пути Энтнера Дудорова. Этот путь отличается у галобактерий тем, что глю-
коза без фосфорилирования окисляется в глюконовую кислоту. Последняя превращается в 2-кето-3-дезоксиглюконовую кислоту, которая расщепляется на два С3-фрагмента: пировиноградную кислоту и глицериновый альдегид. Из глицеринового альдегида в результате нескольких ферментативных реакций также образуется ПВК. Дальнейшее окисление происходит в замкнутом цикле Кребса.
 Основной способ получения энергии экстремальными галофилами аэробное дыхание. Использование световой энергии для создания трансмембр. градиента протонов происходит с участ. бактериородопсина и не связано с переносом электронов по цепи переносчиков. Шиффово основание в темноте находится в протонированной форме. Поглощение кванта света бактериородопсином вызывает изменение конформации ретиналя и приводит к отщеплению Н+ от Шиффова основания. Протон, переходит во внеклеточное пространство, а Н+, протонирующий Шиффово основание, поглощается из цитоплазмы.
Основной способ получения энергии экстремальными галофилами аэробное дыхание. Использование световой энергии для создания трансмембр. градиента протонов происходит с участ. бактериородопсина и не связано с переносом электронов по цепи переносчиков. Шиффово основание в темноте находится в протонированной форме. Поглощение кванта света бактериородопсином вызывает изменение конформации ретиналя и приводит к отщеплению Н+ от Шиффова основания. Протон, переходит во внеклеточное пространство, а Н+, протонирующий Шиффово основание, поглощается из цитоплазмы.
В результате работы циклического механизма, получившего название бактериородопсиновой протонной помпы, при освещении по разные стороны мембраны возникает градиент концентрации Н+, достигающий 200 мВ. Разрядка протонного градиента с помощью Н+-АТФ- синтазы приводит к синтезу АТФ.
Поможем написать любую работу на аналогичную тему