Биосинтез жиров состоит из трех этапов: синтеза глицерина, высших жирных кислот и образования триацилглицеринов.
Синтез глицерина осуществляется из промежуточного продукта гликолиза и пентозофосфатного цикла - дигидроксиацетонфосфата, который глицерофосфатдегидрогеназой восстанавливается в глицерол-З-(Р).
СН2ОН СН2 ОН
ô + НАД· Н2 ô
![]() С = О СНОН + НАД+
С = О СНОН + НАД+
![]()
![]() ô ô
ô ô
СН2 - О- Р СН2 - О- Р
Дигидрокси- Глицерол-3-фосфат
ацетонфосфат
Синтез насыщенных и мононенасыщенных жирных кислот происходит в цитоплазме клеток. Некоторое количество жирных кислот синтезируется в мембранах эндоплазматической сети путём удлинения КоА-производных полиненасыщенных жирных кислот. Удлинение молекул жирных кислот происходит и в митохондриях. Особенно активно синтез протекает в клетках жировой ткани, молочной железы и печени.
Сырьём для биосинтеза, у животных с однокамерным желудком, служит ацетил-КоА, который образуется из избыточной глюкозы пищи, не использованной на энергетические цели, а также из аминокислот, не использованных для обновления белков; у жвачных животных - из низкомолекулярных жирных кислот: уксусной, пропионовой, масляной и др., поступающих из преджелудков (См. Переваривание углеводов). Восстановителем в синтезе служит НАДФ×Н2, -образующийся в пентозофосфатном цикле. Другим источником НАДФ×Н2 служит окисление малата до пирувата и СО2 малатдегидрогеназой (декарбоксилирующей оксалоацетат) КФ:I.I.I.40.:
СООН СООН
ô ½ СООН
Н - С- ОН + НАДФ С = О ½
![]()
![]() ½ ½ С = О
½ ½ С = О
СН2 - НАДФ · Н2 СН2 - СО2 ½
½ ½ СН3
СООН СООН
Малат Оксалоацетат Пируват
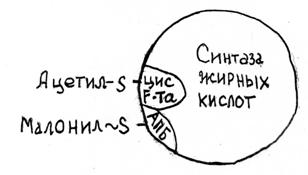
Биосинтез жирных кислот катализирует синтетазная система, состоящая из 7 ферментов объединенных в комплекс М.м.400 тыс. Конечный продукт этого процесса - пальмитиновая кислота -источник всех необходимых насыщенных и мононенасыщенных жирных кислот млекопитающих.
Суммарно процесс биосинтеза пальмитата выглядит так:
Ацетил-КоА + 7 Малонил-КоА + 14 НАДФ×Н2 ® CH3-(CH2)14-COOН + 7 CO2 + + 8 HS-KoA + 14 HAДФ+ + 6 Н2О
Активную роль в биосинтезе играет сложный ацилпереносящий белок (АПБ-SH) М.м. 9000, простетической группой которого является фосфопантотеновая кислота и тиоэтиламин с сульфгидрильной (-SН) группой, как в KoA-SH. Пантотеноая кислота и тиоэтиламин АПБ служат "рукой", переносящей растущую цепь жирной кислоты в комплексе от одного активного центра к другому.
Процесс начинается с образования малонил-KoA из ацетил-КоА и бикарбоната:
Н3С-СО~КоА + НСО-3
+ АТФ ![]() НООС-СН2-СО~SКоА + АДФ + Фн Малонил-КоА (l)
НООС-СН2-СО~SКоА + АДФ + Фн Малонил-КоА (l)
Фермент ацетил-КоА-карбоксилаза активируется цитратом и a-кетоглутаратом. Ацетил-КоА для синтеза, жирных кислот поступает из матрикса митохондрий в форме цитрата, который после выхода в цитоплазму расщепляется ATФ-зависимым ферментом на ацетил-КоА и оксалоацетат.
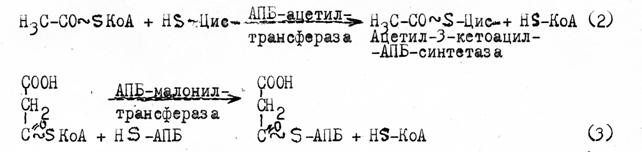 |
Далее ацетил-КоА передает остаток ацетила на НS-группу радикала цистеина фермента 3-кетоацил-АПБ-синтетазы (2) остаток малонила переносится с малонил-КоА на HS-АПБ (3).
 Затем ацетил-S-цис-фермента и малонил-S-АПБ конденсируются, с предварительным декарбоксилированием -СООН группы малонил-S-АПБ. Отщепление СО2 от малонильной группы резко повышает реакционную способность оставшегося фрагмента для взаимодействия с ацетильной группой.
Затем ацетил-S-цис-фермента и малонил-S-АПБ конденсируются, с предварительным декарбоксилированием -СООН группы малонил-S-АПБ. Отщепление СО2 от малонильной группы резко повышает реакционную способность оставшегося фрагмента для взаимодействия с ацетильной группой.
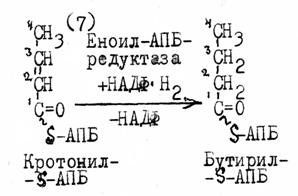
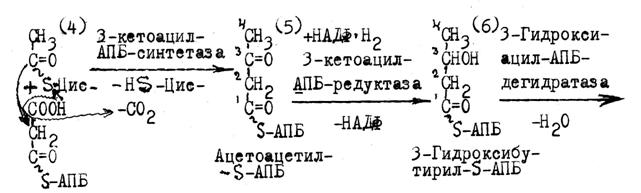
Следующие за 4-й реакцией восстановление, дегидратация и второе восстановление приводят к образованию бутирил-S-АПБ с С4 цепочкой, которая перебрасывается на HS-группу Цистеина фермента. На HS—АПБ переносится вторая молекула малонила. В каждом новом обороте цикла присоединения малонил-S-АПБ (р.4-7) растущая цепочка будет удлиняться на два углеродных атома, до образования пальмитиновой кислоты.
Пальмитиновая и стеариновая кислоты, подвергаясь анаэробному или аэробному окислению превращаются в мононенасыщснные, например, олеиновую.
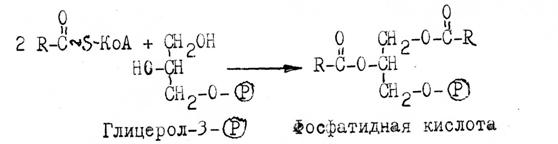 |
Синтез триацилглицеринов происходит главным образом в печени и жировой ткани из КоА-производных жирных кислот и глицерол-3-(Р) через образование фосфатидной кислоты:
Активирование глицерина катализирует глицеролкиназа за счет АТФ. Глицерол-3-Ф (может образоваться и при восстановлении дигидроксиацетон-(Ф) (См. синтез глицерина).
Гидролиз фосфатидной кислоты фосфатазой приводит к образованию 1,2-диацилглицерина, ацилирование которого молекулой ацил-КоА завершает синтез специфичного триацилглицерина.
Биосинтез мембранных глицерофосфолипидов локализован главным образом в эндоплазматической сети клетки. Предшественниками для синтеза являются фосфатидная кислота и активированные азотистые основания. Активирование, например этаноламина, осуществляется так: вначале йод действием этаноламинкиназы образуется фосфоэтаноламин (Этанол-амин+АТФ®фосфоэтаноламин+АДФ).Затем фосфоэтаноламинцитидилтранс-фераза образует цитидиндифосфатэтаноламин (фосфоэтаноламин+ЦТФ ® ЦДФ-этаноламин + Пирофосфат).
Фосфатидная кислота предварительно гидролизуется фосфотазой, с образованием диацилглицерина (фосфатидная кислота + Н2О ® диацилглицерин + Фн). Присоединение полярной группы к диацилглицерину катализирует этаноламинфосфотрансфераза: Диацилглицерин + ЦДФ-этаноламин ® фосфатидилэтаноламин +ЦДФ (см. строение фосфолипидов).
Глицерофосфолипиды, синтезируемые ферментами эндоплазматического ретикулума, не запасаются, а встраиваются в основном в липидный бислой ретикулума. Мембраны ретикулума служат предшественниками мембран аппарата Гольджи, от которого постоянно отшнуровываются мембранные пузырьки, в которых продукты секреции транспортируются к плазматической мембране и часто сливаются с нею (См. Строение клетки). Глицерофосфолипиды могут переноситься из ретикулума в митохондрии и др. органеллы транспортными белками, для замены "изношенных" молекул полярных липидов мембран.
Биосинтез холестерина и стеридов осуществляется из ацетил-КоА, главным образом в тканях печени, головного мозга и надпочечников.
Синтез холестерина насчитывает более 35 реакций. Исследования с ацетатом меченым 14C показали, что процесс имеет три основных стадии: первая - превращение ацетилов в мевалоновую кислоту, вторая - образование из мевалоновой кислоты сквалена, третья - циклизация сквалена в холестерин.
Стериды синтезируются из поступающих с кормами и синтезируемых в клетках холестерина и жирных кислот. Процесс начинается с активирования жирной кислоты путем образования ацил –KoA производного, затем происходит этерификация спиртовой группы холестерина активным остатком жирной кислоты (см. строение стероидов)
Холестерин характеризуется высокой степенью обмена. В течение суток 20 % холестерина (с радиоактивной меткой) окисляется до СО2 и Н2О.
Поможем написать любую работу на аналогичную тему