1. Окисление глюкозы в клетках.
2. Метаболизм пировиноградной кислоты.
3. Схематичное представление полного окисления молекулы глюкозы.
Углеводы наряду с белками и липидами являются важнейшими биологическими соединениями. В организме животных имеются несколько десятков разных моносахаридов и много различных олиго- и полисахаридов. Функции углеводов заключаются в том, что они, прежде всего, служат источником энергии. За счет их окисления обеспечивается около половины всей потребности животного в энергии, при этом главная роль принадлежит глюкозе и гликогену. Углеводы входят в состав структурно-функциональных компонентов клеток. Из углеводов могут синтезироваться соединения других классов, в частности липиды и некоторые аминокислоты. В организме животных и человека наиболее распространенным углеводом является глюкоза.
Глюкоза – постоянная составная часть крови. Несмотря на периодическое поступление и постоянное изымание клетками, уровень глюкозы в крови поддерживается на определенном постоянном, характерном для каждого вида животного уровне. У человека содержание глюкозы в крови колеблется от 80 до 120 мг%. Концентрация глюкозы в крови у различных видов с/х животных составляет, мг%:
лошади 75-95 свиньи 80-100
коровы 40-60 кролики 100-200
В поддержании постоянства уровня глюкозы в крови участвуют множество органов и тканей, физиологических и биохимических процессов. Центральную роль в этом процессе играют гликогенная функция печени и глюконеогенез.
Избыточное количество глюкозы, поступающего из желудочно-кишечного тракта, в клетках печени превращается в гликоген. Реакции синтеза гликогена катализируются 1,4- и 1,6-глюкозилтрансферазами. Последнюю также называют гликогенветвящийся ферментом. В условиях недостаточного поступления глюкозы в кровь или снижения её уровня, гликоген печени расщепляется путем фосфорилаза и служит источником глюкозы.
Гликоген синтезируется во всех клетках организма. Особенно много его в клетках лимфоидной ткани и мышечной ткани, где служит легко мобилизуемым источником энергии.
Глюконеогенез – образование или синтез глюкозы из продуктов неуглеводистого характера. Это может быть метаболиты липидного и белкового обмена, а также другие соединения, такие как пируват, молочная кислота и др.
Углеводный обмен в организме животных и человека складывается из следующих основных процессов:
- расщепление в желудочно-кишечном тракте полисахаридов и дисахаридов пищи, всасывание моносахаридов из кишечника в кровь;
- синтез и распад гликогена в тканях, прежде всего в печени;
- анаэробное и аэробное окисление глюкозы.
- взаимопревращение гексоз;
- метаболизм пирувата. Этот процесс выходит за рамки углеводного обмена, однако может рассматриваться как промежуточная его стадия.
Окисление глюкозы
Окисление глюкозы, как все процессы окисления в целом, бывает аэробное и анаэробное.
Аэробное окисление или прямое окисление, или петозофосфатный путь окисления глюкозы по биоэнергетике не является главным. Основное его предназначение в снабжении клетки с фосфорилированными пентозами, в создании условий для взаимопревращений гексоз, а также в генерации восстановителей типа НАДФН.
Анаэробное окисление глюкозы – гликолиз.
Гликолиз – центральный путь катаболизма глюкозы. Гликолиз – биохимический процесс, в ходе которого молекула глюкозы в десяти последовательных реакциях расщепляется до двух молекул пировиноградной кислоты (пируват).
Условно, гликолиз можно рассматривать в двух стадиях по пять реакций в каждой. Если первую стадию можно
назвать подготовительной, то вторую – энергетической.
Первая стадия:
1. Фосфорилирование глюкозы. Фермент - гексокиназа. Расходуется АТФ, продукт реакции – глюкоза-6-фосфат;
Фермент гексокиназа способен катализировать фосфорилирование не только D-глюкозы, но и других гексоз, в частности D-фруктозы, D-маннозы и т.д. В печени, кроме гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование только D-глюкозы.
2. Изомеризация глюкоза-6-фосфат в фруктоза-6-фосфат. Фермент – фосфоглюкоизомераза.
3. Фосфорилирование фруктозы-6-фосфат с образованием фруктоза-1,6-дифосфат. Фермент - фосфофруктокиназа, расходуется АТФ.
Данная реакция практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза и определяет скорость гликолиза в целом.
4. Альдолазная реакция. Фермент – альдолаза (углерод – углерод лиаза). В целом, можно сказать, что предыдущие реакции служат для подготовки данной реакции. Фруктоза-1,6-дифосфат расщепляется на две фосфотриозы – диоксиацетонфосфат и фосфоглицериновый альдегид.
5. Изомеризация триозофосфатов. Катализируется триозофосфатизомеразой.
Равновесие данной реакции сдвинуто в сторону образования диоксиацетонфосфата. Однако из-за того, что в последующие реакции гликолиза вовлекается только глицеральдегид-3-фосфат, по мере его потребления диоксиацетонфосфат превращается в глицеральдегид-3-фосфат.
Таким образом, в первой стадии, в реакциях фосфорилирования, расходуется 2 молекулы АТФ и образуются триозофосфаты - 2 молекулы 3-фосфоглицеральдегида, которые вовлекаются в окисление во второй стадии.
Вторая стадия:
1. Окислительное фосфорилирование глицеральдегид-3-фосфата. Фермент НАД содержащая дегидрогеназа, участвует неорганический фосфат, продукт реакции - макроэргическое соединение 1,3-дифосфоглицерат. Выделяется НАДН2.
Значение этой реакции в том, что энергия, высвобождающаяся при окислении фосфоглицеральдегида, запасается в макроэргической фосфатной связи дифосфоглицерата.
2. Реакция является логическим продолжением предыдущей. Катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфоглицериновой кислоты (3-фосфоглицерат).
3. Внутримолекулярный перенос фосфатной группы, и превращение 3-фосфоглицерата в 2-фосфоглицерат. Фермент фосфоглюкомутаза.
4. Реакция дегидратации. 2-фосфоглицерат в результате отщепления молекулы воды превращается в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической.
5. Перенос макроэргической фосфатной группы на АДФ. Фермент – пируваткиназа, продукт – пируват и выделяется АТФ.
Во второй стадии образуются 2 молекулы пируват, выделяются 2 НАДН2 и 4 АТФ.
Суммарное уравнение гликолиза:
Глюкоза ® 2 пируват + 2НАДН2 + 2АТФ
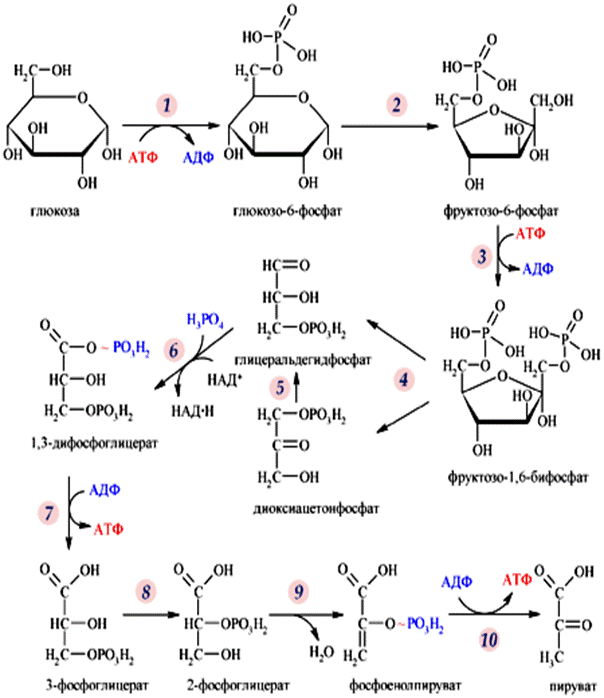
Рис. 18. Химизм гликолиза.
В гликолиз могут вовлекаться и другие углеводы, только моносахариды при этом должны превращаться в глюкозу. Если гликолиз начинается с предварительного расщепления гликогена, т.е. источником глюкозы является гликоген, то процесс называется – гликогенолиз.
Метаболизм пирувата.
Пируват – конечный продукт гликолиза. В организме высших животных и человека в зависимости от условий и состояния клеток подвергается дальнейшему превращению двумя основными способами.
1. В условиях кислородной недостаточности пируват превращается в молочную кислоту (лактат). Гликолиз, с последующим превращением пирувата в лактат, называется молочнокислое брожение. В анаэробных условиях, когда кислород не успевает доставляться к тканям, например, клеткам мышечной ткани при больших физических нагрузках, данный процесс служит одним из важнейших источников АТФ.
Брожение – общий термин, означающий анаэробный распад органических молекул. В основе брожения углеводов лежит гликолиз. По конечному продукту выделяют различные виды брожения, и все они эволюционно развивались как способ добывания живыми клетками энергии из органических молекул.
Молочнокислое брожение, например, лежит в основе жизнедеятельности молочнокислых бактерий.
У некоторых микроорганизмов: дрожжей рода Saccharomyces, бактерий различных таксономических групп (Erwinia amylovora, Zymomonas mobilis), а также у мезофильных и термофильных бактерий (Lactococcus lactis, Clostridium thermohydrosulfuri-cum) пируват превращается в этиловый спирт – спиртовое брожение.
Различают также пропионовокислое, маслянокислое и др. виды брожения.
2. В аэробных условиях пировиноградная кислота подвергается окислительному декарбоксилированию в присутствии кофермента А и превращается в ацетил-КоА. Дальнейшая судьба ацетильных групп весьма предсказуема – окисление в цикле Кребса с последующим переносом электронов в дыхательной цепи.
Схематичное представление полного окисления
молекулы глюкозы.
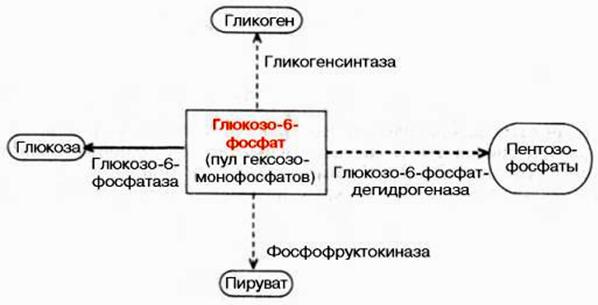 Любые химические превращения глюкозы в клетках животных и человека начинается с её фосфорилирования под действием ферментов гексо- или глюкокиназ с образованием глюкоза-6-фосфат. 6-фосфоглюкоза в дальнейшем может использоваться в трех основных направлениях.
Любые химические превращения глюкозы в клетках животных и человека начинается с её фосфорилирования под действием ферментов гексо- или глюкокиназ с образованием глюкоза-6-фосфат. 6-фосфоглюкоза в дальнейшем может использоваться в трех основных направлениях.
Рис. 19. Основные пути превращений глюкозы
Во-первых, синтез гликогена. Первая реакция на этом пути - превращение глюкозы-6-фосфат в глюкозу-1-фосфат, катализируется фосфоглюкомутазой.
Во-вторых, аэробное окисление или пентозофосфатный путь превращения. Первая реакция этого пути превращений - прямое окисление 6-фосфоглюкозы с образованием 6-фосфоглюконат.
В третьих, вовлечение глюкозы в гликолиз.
Полное окисление глюкозы (биологическое окисление) до СО2 и Н2О можно рассматривать в трех стадиях.
Первой стадия - гликолиз.
Глюкоза => 2 Пируват + 2НАДН2 + 2 АТФ;
Вторая или промежуточная стадия - окислительное декарбоксилирование пирувата:
2 Пируват => 2Ацетил-КоА + 2НАДН2
Заключительная стадия - окисление 2 молекул ацетил-КоА в цикле Кребса с последующим переносом электронов в дыхательной цепи.
2Ацетил-КоА => 2 АТФ + 6НАДН2 + 2 ФАДН2 = 24 АТФ (см.выше).
Таким образом, суммарный выход АТФ при полном окислении одной молекулы глюкозы - 38 молекул.
Поможем написать любую работу на аналогичную тему