1. Окисление высших жирных кислот и глицерина.
2. Энергетический баланс окисления ВЖК и глицерина.
3. Метаболизм кетоновых тел.
4. Биосинтез жиров и фосфолипидов.
Липиды - обширная группа биоорганических соединений и поэтому не имеют единой химической характеристики. Однако, в известной мере их можно рассматривать как класс органических соединений, большинство из которых принадлежат сложным эфирам многоатомных или специфически построенных спиртов с высшими жирными кислотами. Липиды характеризуются также различной растворимостью в органических растворителях и, как правило, нерастворимы в воде.
В зависимости от химического состава липиды подразделяют на простые и сложные. Простые липиды включают вещества, молекулы которых состоят только из остатков жирных кислот и спиртов. К ним относят жиры (триглицириды) – сложные эфиры высших жирных кислот и трехатомного спирта – глицирина; воска – сложные эфиры высших жирных кислот и высших спиртов; стериды – сложные эфиры высших жирных кислот и полициклических спиртов – стеролов.
Сложные липиды имеют многокомпонентные молекулы, компоненты которых соединены химическими связями различного типа. К ним принадлежат: фосфолипиды, состоящие из остатков высших жирных кислот, глицерина или других многоатомных спиртов, фосфорной кислоты и азотистых оснований той или иной природы; гликолипиды, включающие в свой состав наряду с многоатомным спиртом и высшей жирной кислотой также углеводы. Однако следует отметить, что существуют и другие методы классификации липидов.
Липиды играют важную роль в процессах жизнедеятельности организма. Они обладают способностью образовывать со многими высокомолекулярными соединениями комплексы. В виде таких комплексов, особенно с белками, липиды входят в состав мембранных структур. Образуя структурный матрикс в виде двойных липидных слоёв, липиды являются основой любой биологической мембраны, влияют на их проницаемость, участвуют в передаче нервного импульса, создании межклеточных контактов. Благодаря особенностям и участию мембранного аппарата, в клетке реализуются такие важные биологические функции липидов, как регуляция деятельности ряда гормонов и активности ферментов (сейчас известно несколько сотен липидзависимых ферментов); влияние на процессы транспорта метаболитов и макромолекул, контроль реакций биологического окисления и энергетического обмена и др. Незаменима роль липидов, особенно жиров, в обеспечении организма энергией. При окислении 1моля жира в организме выделяется 9,3 ккал. энергии. Естественно, что при окислении липидов возникают метаболиты, широко вовлекаемые в биосинтез других соединений.
Промежуточный обмен жиров включает следующие основные процессы:
- расщепление триглицеридов в тканях с образованием высших жирных кислот и глицерина;
- окисление ВЖК и глицерина;
- образование и расщепление кетоновых тел;
- биосинтез жиров, фосфоглицеридов и др. классов липидов.
Окисление высших жирных кислот
Центральным биохимическим процессом в катаболизме липидов является окисление высших жирных кислот. Различают несколько видов окисления ВЖК, среди которых центральное место занимает β-окисление. β-окисление происходит в матриксе митохондрий клеток, практически всех тканей организма. Наиболее интенсивно протекает в клетках печени, а в мозговой ткани скорость окисления жирных кислот весьма незначительна. Объясняется это тем, что основным источником энергии в мозговой ткани служит глюкоза. Впервые выдвинул гипотезу β-окисления жирных кислот в 1904 г. Франц Кнооп. Большой вклад в расшифровку механизмов β-окисления внесли также Альберт Ленинджер и Феодор Линен (спираль Линена).
β-окисление - это расщепление длинной углеводородной цепи ВЖК до молекул ацетил-КоА в циклично повторяемых, последовательных 4-х реакциях.
Свободная жирная кислота является метаболически инертной и не может подвергаться никаким биохимическим превращениям. Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ и коэнзима A. Реакция катализируется ферментом ацил-КоА-синтетазой и в результате образуется ацил-КоА - активная форма жирной кислоты.
Переносчиком активированных жирных кислот
с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин.
За каждый цикл процесса молекула жирной кислоты укорачивается на 2 атома углерода. Как побочные продукты выделяются также по одной молекуле НАДН2 и ФАДН2.
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Поэтому любая такая кислота, от которой отщепляется по паре углеродных атомов, в конечном итоге проходит через стадию масляной кислоты (четырехуглеродный фрагмент – бутирил-КоА). После очередного цикла β-окисления бутирил-КоА распадается на две молекулы ацетил-КоА.
Таким образом, при β-окислении любой ВЖК на последней стадии выделяются 2 молекулы ацетил-КоА. Следовательно, количество этапов (циклов) всегда меньше чем количество образовавшихся ацетил-КоА на одного. Это важно знать в расчетах энергетического баланса процесса.
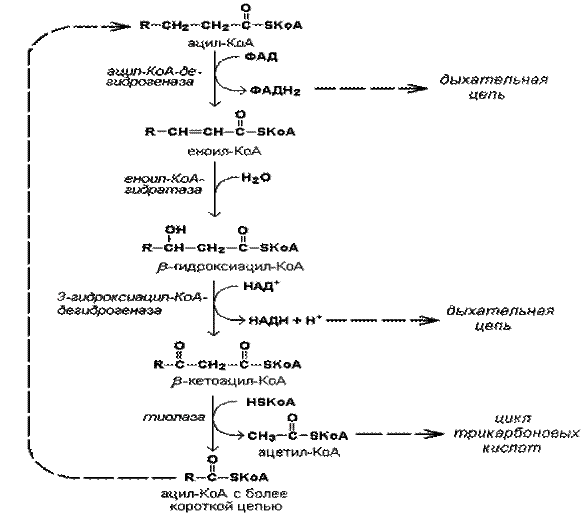
Рис. 20. Химизм β-окисления ВЖК
Энергетический баланс β-окисления
Валовое уравнение каждого цикла можно выразить следующей схематичной формулой:
ВЖК -> НАДН2 + ФАДН2 + ацетил-КоА + ВЖК, укороченная на 2 атома углерода.
В расчетах энергетического баланса биохимических процессов необходимо уметь выделять основные стадии и знать валовые уравнения каждого этапа.
При полном окислении жирных кислот начальной стадией процесса является β-окисление, а заключительной – окисление ацетил-КоА в цикле Кребса с последующим переносом электронов и окислительным фосфорилированием в дыхательной цепи.
Например, необходимо вычислить суммарный выход АТФ при полном окислении одной молекулы миристиновой кислоты.
Углеводородная цепь миристиновой кислоты состоит из 14 углеродов. Следовательно, расщепление её на 7 молекул ацетил-КоА в β-окислении произойдет за 6 циклов. Валовое уравнение:
Миристиновая кислота —> 6НАДН2 + 6ФАДН2 + 7ацетил-КоА.
6НАДН2 = 6x3=18 АТФ;
6ФАДН2 = 6x2= 12 АТФ;
7ацетил-КоА = 7 х 12 = 84 АТФ (см. окисление в цикле Кребса);
В сумме, при полном окислении одной молекулы миристиновой кислоты образуется 114 АТФ.
Окисление глицерина
Глицерин, образовавшийся при гидролизе жиров и других глицеридов, может подвергаться различным химическим превращениям. Любые превращения глицерина начинаются с его фосфорилирования и образования фосфоглицерина. Фермент – глицеролкиназа, расходуется АТФ. 3-фосфоглицерин с участием НАД содержащей дегидрогеназы подвергается окислению и образуется 3-фосфоглицериновый альдегид – промежуточный продукт гликолиза. В связи с этим, дальнейшая судьба фосфоглицеринового альдегида весьма предсказуема. Конечно, в зависимости от условий и потребностей организма он может использоваться разными путями, но с точки зрения биоэнергетики путь один – окисление по схеме гликолиза.
Метаболизм кетоновых тел
К кетоновым телам относят три соединения:
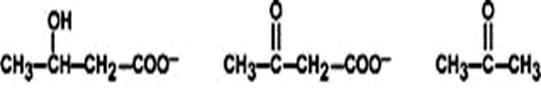
3-оксибутират ацетоацетат ацетон
В крови здорового человека кетоновые тела содержатся лишь в очень небольших количествах (0,3–0,2 ммоль/л). При некоторых патологических состояниях, при голодании концентрация кетоновых тел в сыворотке крови резко увеличивается и может достигать 16–20 ммоль/л.
Кетоновые тела образуются в печени. Стимулом для образования кетоновых тел служит поступление большого количества жирных кислот в печень. При состояниях активирующих липолиз в жировой ткани, не менее 30% жирных кислот задерживаются печенью. Однако, нужно помнить, что синтез кетоновых тел идет постоянно. Считается доказанным ведущая роль кетоновых тел в поддержании энергетического гомеостаза. Кетоновые тела – своего рода поставщики топлива для мышц, мозга, почек. Сердечная мышца, кора надпочечников используют как источник энергии ацетоацетат гораздо больше, чем глюкозу в физиологических условиях.
Механизм синтеза кетоновых тел может идти двумя путями, в частности с конденсацией двух или трех молекул ацетил-КоА. Центральное место, как в механизме синтеза, так и в механизме распада кетоновых тел занимает ацетоацетат. Ацетоацетат может восстанавливаться до 3-оксибутират или декарбоксилироваться с образованием ацетона. Кетоновые тела поступают в кровь и разносятся по тканям. Ацетон, как летучее вещество, легко удаляется выдыхаемым воздухом или потом. В целом, можно предположить, что его образование происходит спонтанно и никакой физиологической роли не играет.
Реакции утилизации кетоновых тел примерно совпадают с обратным направлением реакций синтеза. Ацетоацетат активизируется в митохондриях клеток периферических тканей и подвергается расщеплению по механизму тиолазной реакции β-окисления. Образуется две молекулы ацетил-КоА, которые обеспечивают, при полном окислении, выход 24 АТФ. Таким образом, образование и распад кетоновых тел в организме животных может считаться одним из основных способов перераспределения энергии.
Биосинтез жиров и фосфолипидов.
Синтез триглицеридов происходит из глицерина и жирных кислот, через образование 3-фосфоглицерина как промежуточного соединения.
В тканях, где активность фермента глицеролкиназы высока например, в почках, глицерин фосфорилируется за счет АТФ с образованием глицерол-3-фосфата. В жировой ткани, в мышцах вследствие низкой активности этого фермента, образование глицерол-3-фосфата сопряжено с гликолизом и происходит за счет восстановления диоксиацетонфосфата.
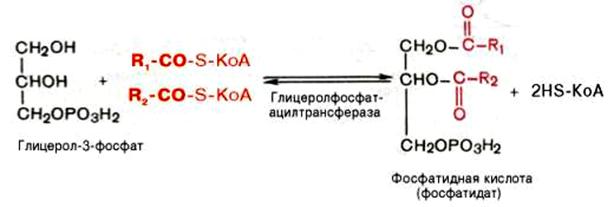 Образовавшийся тем или иным путем 3-фосфоглицерин ацилируется двумя молекулами ацил- КоА. В результате образуется фосфатидная кислота:
Образовавшийся тем или иным путем 3-фосфоглицерин ацилируется двумя молекулами ацил- КоА. В результате образуется фосфатидная кислота:
Фосфатидная кислота является общим промежуточным продуктом, как при синтезе жиров, так и фосфоглицеридов.
При синтезе жиров фосфатидная кислота дефосфорилируется (щелочная фосфотаза) и происходит дальнейшее ацилирование.
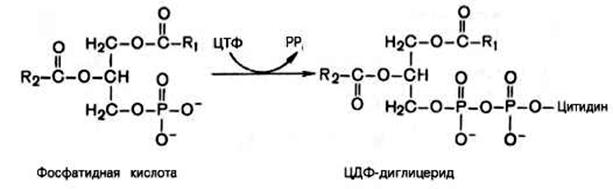 Синтез фосфоглицеридов начинается с взаимодействия фосфатидной кислоты с ЦТФ. Образуется соединение – диглицерид-ЦДФ, являющийся общим продуктом при синтезе всех фосфолипидов.
Синтез фосфоглицеридов начинается с взаимодействия фосфатидной кислоты с ЦТФ. Образуется соединение – диглицерид-ЦДФ, являющийся общим продуктом при синтезе всех фосфолипидов.
Затем происходит перенос на фосфатидильный остаток серина с образованием фосфатидилсерина – предшественника фосфатидилэтаноламина (кефалина), фосфатидилхолина (лейцитина).
Поможем написать любую работу на аналогичную тему