Глюкоза в клетках человека, животных, растений и многих бактерий окисляется в две стадии: анаэробную и аэробную.
Гликолиз - анаэробная стадия окисления глюкозы в тканях человека и животных до двух молекул лактата (молочной кислоты). При этом освобождающаяся энергия аккумулируется в молекулах АТФ. Гликолиз протекает в цитоплазме, состоит из 11 реакций и условно делится на две фазы. Первая фаза - активирование глюкозы (пусковая реакция, тормозится её продуктом - глюкозо-6-фосфатом) и подготовка к расщеплению на две фосфотрнозы.(реакции 1-3); расщепление фруктозо-1,6-дифосфата и изомеризация дигидроксиацетонфосфата в глицеральдегид-3-фосфат (реакции 4,5). Вторая фаза - окисление двух молекул глицеральдегид-3-фосфата до двух молекул лактата и аккумуляция освобождающейся энергии в сопряженном процессе субстратного фосфорилирования АДФ с образованием АТФ (реакции 6-11).
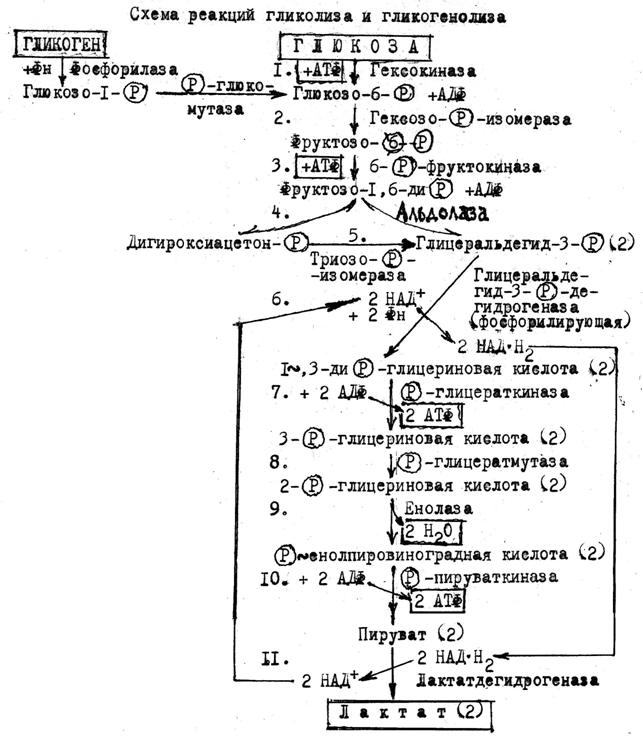
*В рамки помещены исходные субстраты и конечные продукты гликолиза; цифрами с точкой помечены номера реакций; цифрой 2 без скобок и в скобках обозначено число молекул.
В анаэробных условиях НАД+ выполняет роль кофермента двух дегидрогеназ и переносчика от глицеральдегид-3-фосфата (реакция 6) на пируват (реакция 11).
Значение гликолиза особенно велико для тканей с ограниченным доступом кислорода и испытывающих периодически резкое возрастание потребления АТФ. Например, в работающих мышцах скорость потребления АТФ может возрасти более чем в 100 paз. В таких условиях мышечная ткань получает основное количество АТФ за счет гликолиза.
При продолжительных интенсивных мышечных нагрузках лактат, накапливаясь в клетках мышц до определенного уровня, выбрасывается в межклеточные пространства, обуславливая чувство усталости. Во время отдыха, в аэробных условиях, лактат окисляется в пируват. До 20 % пирувата окисляется до СО2 и Н2О для получения энергии, около 80 % - идет на ресинтез глюкозы и гликогена.
Гликолиз может начинаться либо с фосфорилирования свободной глюкозы, либо с фосфоролиза гликогена. В скелетных мышцах оба пути выражены в равной степени, а в мышце сердца и головном мозгу преобладает фосфорилирование глюкозы.
Брожения - это окисление углеводов или др. органических веществ некоторыми микроорганизмами для получения энергии в анаэробных условиях. Изучено несколько типов бржений, их химизм до образования пирувата одинаков с гликолизом. Различаются брожения реакциями преобразования пирувата в конечные продукты характерные для микроорганизмов, осуществляющих данный тип брожения.
Молочнокислое брожение - это процесс получения энергии молочнокислыми бактериями. По характеру брожения различают две группы молочнокислых бактерий: гомоферментативные и гетероферментативные, осущетвляющих соответствующий тип брожения.
Гомоферментативное молочнокислое брожение - это процесс осуществляемый гомоферментативными (однотипно-бродящими) бактериями, которые образуют в основном молочную кислоту и очень мало побочных продуктов. Они окисляют углеводы молока по пути гликолиза в анаэробных условиях, где пируват служит акцептором от НАД×Н2 и восстанавливается в конечный продукт брожения - лактат (реакция 11). Суммарно этот тип брожения можно записать так:
C6Н12О6![]() 2СН3-СНОН-СООН+2АТФ
2СН3-СНОН-СООН+2АТФ
Лактат, накапливаясь до рН 4,6, вызывает скисание молока. Этот же процесс лежит в основе квашения капусты, огурцов, помидор и др. продуктов растительного происхождения, силосования кормов для животных.
Гетероферментативное молочнокислое брожение - это процесс осуществляемый гетероферментативными (разнотипно- бродящими) бактериями. Наряду с молочной кислотой они образуют значительное количество других веществ - этанола, СО2, некоторые ещё уксусной кислоты.
Окисление углеводов молока гетероферментативными бактериями обуславливается своеобразием ферментной системы гетероферментативного молочнокислого брожения в которой нет фермента альдолазы. После фосфорилирования гексоза окисляется дегидрогеназой и декарбоксилируется , превращаясь в пентозофосфат. Последний расщепляется на глицеральдегид-3-фосфат и ацетилфосфат. Глицеральдегид -3-фосфат, как и у гомоферментативных молочнокислых бактерий, окисляется до пирувата, который затем восстанавливается в молочную, а НАД×Н2 окисляется в НАД+:
![]()
Ацетилфосфат дефосфорилируется и превращается в уксусную кислоту, частично восстанавливается (через уксусный альдегид) в этанол. Таким оброзом, конечными акцепторами в этом типе брожения служат пируват и уксусный альдегид.
Культуры гетероферментативных молочнокислых бактерий используют в производстве кефира, кумыса, курунги, мацони и др. продуктов.
Спиртовое брожение осуществляется клетками дрожжей для получения энергии в анаэробных условиях. Большинство дрожжей сбраживает моносахариды (главным оброзом гексозы), из дисахаридов - сахарозу и мальтозу по пути гликолиза до образования пирувата. В анаэробных условиях, синтезируемый дрожжами фермент пируватдекарбоксилаза, превращает пируват в уксусный альдегид. Последний ферментом алкогольдегидрогеназой восстанавливается в этанол с окислением НАД×Н2:
![]() Пируват Укс. Альдегид Этанол
Пируват Укс. Альдегид Этанол
Суммарное уравнение спиртового брожения выглядит так:
![]()
Есть микроорганизмы осуществляющие уксуснокислое, пропионовокислое, маслянокислое, лимоннокислое и др. виды брожений. Многие брожения используются в пищевых и др. технологиях, не менее важную роль они выполняют в природе.
Пентозофосфатный путь окисления глюкозы
Наряду с гликолизом, поставляющим энергию, существуют второстепенные пути катаболизма глюкозы, нарабатывающие особые продукты необходимые клеткам. Рассмотрим один из них - пентозофосфатный.
Пентозофосфатный путь имеет две фазы, все реакции протекают в цитоплазме. Первая фаза окислительная: глюкозо-6-фосфат окисляется НАДФ-зависимой глюкозо-6-фосфат дегидрогеназой до 6-фосфоглюконата, который затем подвергается дегидрированию и декарбоксилированию с образованием рибулозо-5-фосфата (кетозы) и ещё одной молекулы НАДФ×H2. Пентозофосфатизомераза превращает рибулозо-5-фосфат в рибозо-5-фосфат (альдозу), который может использоваться на биосинтез рибонуклеотидов и дезоксирибонуклеотидов. В некоторых клетках (например, мышечной ткани) пентозофосфатный путь на этом кончается. Суммарное уравнение первой фазы можно записать так:
Глюкозо-6-фосфат + 2 НАДФ + Н20® Рибозо-5-фосфат + СО2 + 2 НАДФ×Н2
Вторая фаза - неокислительная, в ней происходят структурные взаимопревращения пентозофосфатов с появлением трех-, четырех-, пяти-, шести-, семи- и восьмиуглеродных фосфосахаров, в результате регенерируется глюкозо-6-фосфат и нарабатывается восстановитель в форме HAДФ×H2. Суммарное уравнение процесса имеет следующий вид:
6 Глюкозо-6-фосфат + 7 Н20 + 12 НАДФ ®
5 Глюкозо-6-фосфат + 6 СО2 + 12 HAДФ×Н2 + Фн
Активность пентозного цикла высока в клетках печени, жировой ткани, молочной железы, коры надпочечников, где он нарабатывает значительное количество НАДФ×Н2 для обеспечения процессов биосинтеза жирных кислот, холестерина, стероидов и др. веществ восстановителем.
В плане энергетики (участия в окислительном фосфорилировании) пентозофосфатный путь существенной роли не играет, т.к. протекая в цитоплазме он может переходить в гликолиз. Существует три точки пересечения пентозного пути с гликолизом: на уровне глюкозо-6-фосфата, фруктозо-6-фосфата и глицеральдегид-3-фосфата. То есть, гликолиз и пентозный путь способны вовремя переключаться с одного на другой.
Аэробная стадия дыхания
Клетки человека, животных, растений, многих микроорганизмов при достаточном поступлении кислорода, образующийся при гликолизе пируват окисляют до СО2 и Н2О в аэробной стадии, которую называют клеточным дыханием.
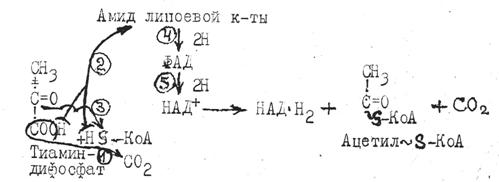 |
В клеточном дыхании, протекающем в митохондриях, различают три стадии. На первой стадии клеточное "топливо": пируват, жирные кислоты окисляются до ацетилкофермента А (ацетил-КоА).
Пируват, образовавшийся в цитоплазме, переносится в митохондрию и подвергается окислительному декарбоксилированию пируватдегидрогеназной ферментной системой, состоящей из трех ферментов и пяти кофакторов (тиаминдифосфат, амид липоевой кислоты, кофермент А, ФАД и НАД+). Процесс протекает в пять этапов завершается образованием макроэргического соединения ацетил-КоА: (активного остатка уксусной кислоты).
Ацетилкофермент А является промежуточным продуктом окисления не только углеводов, но и глицерина, жирных кислот, ряда a-аминокислот. Ацетилкофермент А служит исходным материалом для синтеза многих органических веществ и основным клеточным "топливом" в аэробных условиях.
Цикл трикарбоновых кислот (ЦТК). (Цикл лимонной кислоты, цикл Кребса)
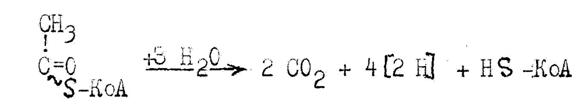 |
ЦПК является стадией аэробного окисления ацетилкофермента А по схеме:
Цикл начинается переносом ацетила с кофермента А на щавелевоуксусную к-ту (ЩУК) или оксалоацетат; их конденсацией и образованием лимонной кислоты ферментом цитратсинтетазой (см. общую схему биологического окисления). В последующих реакциях цикла ацетил окисляется до двух молекул СО2 и четырех пар водорода (4 ): три пары снимаются НАД+-зависимыми, а одна - ФАД-зависимой дегидрогеназой. Завершающая реакция цикла - окисление яблочной кислоты в ЩУК, которая способна вступать в новьй цикл реакций. Фермент НАД+-зависимая изоцитратдегидрогеназа аллостерически активируется АДФ и ингибируется АТф, увеличивая или замедляя (до полной остановки) окисление изолимонной кислоты и скорость последующих реакций ЦТК.
Окислительное декарбоксилирование a-кетоглутаровой кислоты осуществляет a-кетоглутаратдегидрогеназная система, по составу, структуре и функции аналогичная пируватдегидрогеназной системе. Конечный продукт процесса - сукцинил ~S-КоА - высокоэнергетический тиоэфир янтарной кислоты (сукцината).
На следующем этапе сукцинил ~S-КоА утрачивает кофермент А, освобождающаяся при этом энергия аккумулируется в макроэнергетической связи ГТФ:
Сукцинил S-КоА+Фн+ГДФ ® Янтарная кислота+ГТФ+KoA-SH.
Реакция катализируется ферментом сукцинил- КоА- синтетазой и относится к реакциям субстратного фосфорилирования.
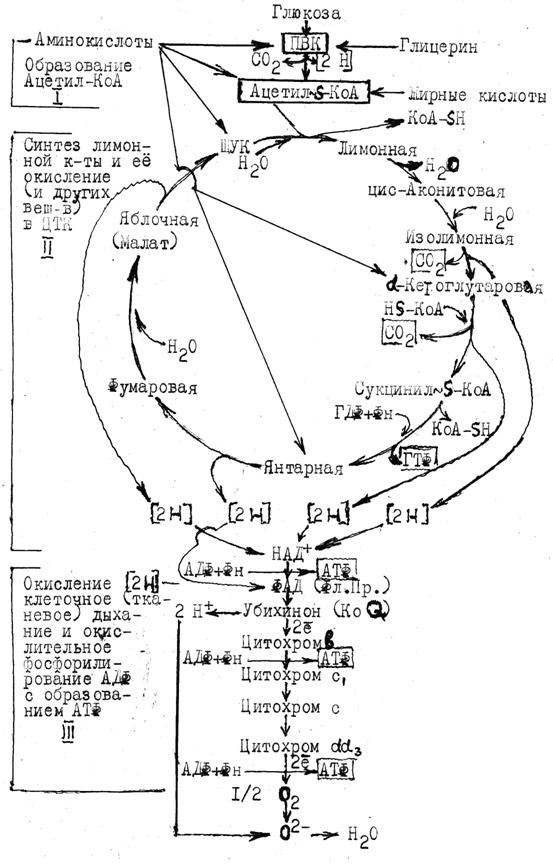 |
Общая схема биологического окисления и стадий дыхания
Многие аминокислоты, подвергаясь реакциям переаминирования и дезаминирования, могут превращаться в метаболиты цикла, например,
щавелевоуксусную, a-кетоглутаровую, янтарную кислоту и, таким образом, вступать в него. Промежуточные продукты цикла, в свою очередь, могут выводиться из него и использоваться для синтеза заменимых аминокислот и др. необходимых веществ.
Поможем написать любую работу на аналогичную тему